Anatomic knowledge of the uterus was slow to accumulate.1,2 Papyrus writings from 2500 B.C. indicate that the ancient Egyptians made a distinction between the vagina and uterus. Because the dead had to be embalmed, dissection was precluded, but prolapse was recognized because it was important to return the uterus into its proper place prior to mummification. Next to the Egyptian papyri in antiquity were Hindu writings in which descriptions of the uterus, tubes, and vagina indicate knowledge gained from dissections. This was probably the earliest description of the fallopian tubes.
There is little information in Greek writings about female anatomy; however, Herophilus (fourth century B.C.), the great anatomist in Alexandria and the originator of scholarly dissection, recorded the different positions of the uterus. Soranus of Ephesus (98-138 A.D.) accurately described the uterus (probably the first to do so), obviously from multiple dissections of cadavers. He recognized that the uterus is not essential for life, acknowledged the presence of leiomyomas, and treated prolapse with pessaries.
Herophilus and Soranus were uncertain about the function of the fallopian tubes, but Galen, Rufus, and Aetisu correctly guessed their function. Galen promoted the practice of bleeding for the treatment of almost every disorder. In his argument that nature prevented disease by discharging excess blood, Galen maintained that women were healthier because their superfluous blood was eliminated by menstruation.3 The writings of Galen (130-200 A.D.) represented the knowledge of medicine for over 1,000 years until the end of the medieval Dark Ages. Galen’s description of the uterus and tubes indicates that he had only seen the horned uteri of animals.
In the 16th century, Berengarius, Vesalius, Eustachius, and Fallopius made significant contributions to the anatomic study of the female genitalia. Berengarius (Giacomo Berengario da Carpi) was the first anatomist to work with an artist. His anatomic text, published in 1514, depicted dissected subjects as if they were still alive.
Gabriele Fallopio (or Fallopius) published his work, Observationes Anatomicae, in Venice in 1561, 1 year before his death from pleurisy at age 40. He provided the first descriptions of the clitoris and the hymen and the first exact descriptions of the ovaries and the tubes. He named the vagina and the placenta and called the tubes the uteri tuba (the trumpet of the uterus), but soon they were known universally as the fallopian tubes. It was his professor and mentor at the University of Padua, however, Andreas Vesalius, who was the first to accurately reveal the presence of the endometrial cavity.
Development of the Müllerian System
The wolffian (mesonephric) and müllerian (paramesonephric) ducts are discrete primordia that temporarily coexist in all embryos during the ambisexual period of development (up to 8 weeks). Thereafter, one type of duct system persists normally and gives rise to special ducts and glands, whereas the other disappears during the third fetal month, except for nonfunctional vestiges.
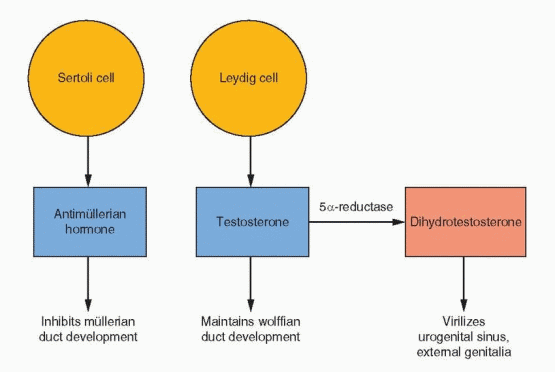
Hormonal control of mammalian somatic sex differentiation was established by the classic experiments of Alfred Jost.4 In Jost’s landmark studies, the active role of male-determining factors, as opposed to the constitutive nature of female differentiation, was defined as the directing feature of sex differentiation. This principle applies not only to the internal ducts but to the gonad, external genitalia, and even the brain. The critical factors in determining which of the duct structures stabilize or regress are the secretions from the testes: AMH (antimüllerian hormone, also known as müllerian-inhibiting substance or müllerian-inhibiting factor) and testosterone.
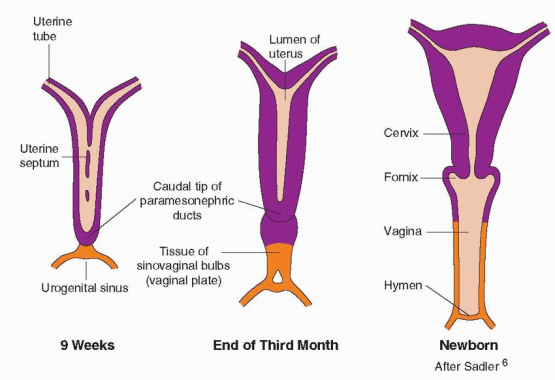
AMH is a member of the transforming growth factor-β family of glycoprotein differentiation factors that include inhibin and activin. The gene for AMH has been mapped to chromosome 19. AMH is synthesized by Sertoli cells soon after testicular differentiation and is responsible for the ipsilateral regression of the müllerian ducts by 8 weeks. Despite its presence in serum up to puberty, lack of regression of the uterus and tubes is the only consistent expression of AMH gene mutations. In the absence of AMH, the fetus will develop fallopian tubes, uterus, and upper vagina from the paramesonephric ducts (the müllerian ducts). This development requires the prior appearance of the mesonephric ducts, and for this reason, abnormalities in development of the tubes, uterus, and upper vagina are associated with abnormalities in the renal system.
The internal genitalia possess the intrinsic tendency to feminize. In the absence of a Y chromosome and a functional testis, the lack of AMH allows retention of the müllerian system and development of fallopian tubes, uterus, and upper vagina. In the absence of testosterone, the wolffian system regresses. In the presence of a normal ovary or the absence of any gonad, müllerian duct development takes place. This process is discussed in greater detail in Chapter 9.
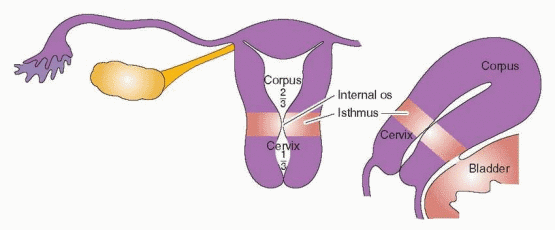
The paramesonephric ducts come into contact in the midline to form a Y-shaped structure, the primordium for the uterus, tubes, and the upper one-third of the vagina.5 The fallopian tubes, uterus, and the upper portion of the vagina are created by the fusion of the müllerian ducts by the tenth week of gestation. Canalization to create the uterine cavity, the cervical canal, and the vagina is complete by the 22nd week of gestation. Under the epithelium lies mesenchymal tissue that will be the origin of the uterine stroma and smooth muscle cells. By the 20th week of pregnancy, the uterine mucosa is fully differentiated into the endometrium.
The endometrium, derived from the mucosal lining of the fused müllerian ducts, is essential for reproduction and may be one of the most complex tissues in the human body. It is always changing, responding to the cyclic patterns of estrogen and progesterone of the ovarian menstrual cycle and to a complex interplay among its own autocrine and paracrine factors.
The Histologic Changes in Endometrium During an Ovulatory Cycle
The sequence of endometrial changes associated with an ovulatory cycle has been carefully studied by Noyes in the human and Bartlemez and Markee in the subhuman primate.7,8,9,10 and 11 From these data a description of menstrual physiology has been developed based on specific anatomic and functional changes within glandular, vascular, and stromal components of the endometrium.12,13 and 14 These changes will be discussed in five phases: (1) the menstrual endometrium, (2) the proliferative phase, (3) the secretory phase, (4) preparation for implantation, and finally, (5) the phase of endometrial breakdown. Although these distinctions are not entirely arbitrary, it must be recalled that the entire process is an integrated evolutionary cycle of endometrial growth and regression, which is repeated some 400 times during the adult life of the human female.
The endometrium can be divided morphologically into an upper two-thirds “functionalis” layer and a lower one-third “basalis” layer. The purpose of the functionalis layer is to prepare for the implantation of the blastocyst; therefore, it is the site of proliferation, secretion, and degeneration. The purpose of the basalis layer is to provide the regenerative endometrium following menstrual loss of the functionalis.15
The Uterine Vasculature
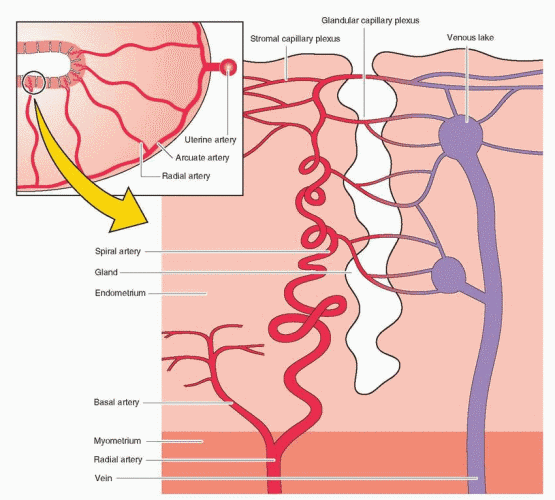
The two uterine arteries that supply the uterus are branches of the internal iliac arteries. At the lower part of the uterus, the uterine artery separates into the vaginal artery and an ascending branch that divides into the arcuate arteries. The arcuate arteries run parallel to the uterine cavity and anastomose with each other, forming a vascular ring around the cavity. Small centrifugal branches (the radial arteries) leave the arcuate vessels, perpendicular to the endometrial cavity, to supply the myometrium. When these arteries enter the endometrium, small branches (the basal arteries) extend laterally to supply the basalis layer. These basal arteries do not demonstrate a response to hormonal changes. The radial arteries continue in the direction of the endometrial surface, now assuming a corkscrew appearance (and now called the spiral arteries), to supply the functionalis layer of the endometrium. It is the spiral artery (an end artery) segment that is very sensitive to hormonal changes. One reason that the functionalis layer is more vulnerable to vascular ischemia is that there are no anastomoses among the spiral arteries. The endometrial glands and the stromal tissue are supplied by capillaries that emerge from the spiral arteries at all levels of the endometrium. The capillaries drain into a venous plexus and eventually into the myometrial arcuate veins and into the uterine veins. This unique vascular architecture is important in allowing a repeated sequence of endometrial growth and desquamation.
The Menstrual Endometrium
The menstrual endometrium is a relatively thin but dense tissue. It is composed of the stable, nonfunctioning basalis component and a variable, but small, amount of residual stratum spongiosum. At menstruation, this latter tissue displays a variety of functional states including disarray and breakage of glands, fragmentation of vessels and stroma with persisting evidence of necrosis, white cell infiltration, and red cell interstitial diapedesis. Even as the remnants of menstrual shedding dominate the overall appearance of this tissue, evidence of repair in all tissue components can be detected. Endometrial regeneration originates in epithelial and stromal stem cells.16 Endometrial epithelial stem cells are located in the base of the endometrial glands and stromal stem cells around blood vessels in the basalis layer.
The menstrual endometrium is a transitional state bridging the more dramatic proliferative and exfoliative phases of the cycle. Its density implies that the shortness of height is not entirely due to desquamation. Collapse of the supporting matrix also contributes significantly to the shallowness. Reticular stains in Rhesus endometrium confirm this “deflated” state. Nevertheless, as much as two-thirds of the functioning endometrium is lost during menstruation. The more rapid the tissue loss, the shorter the duration of flow. Delayed or incomplete shedding is associated with heavier flow and greater blood loss.
DNA synthesis is occurring in those areas of the basalis that have been completely denuded by day 2-3 of the menstrual cycle (the endometrium in the isthmic area, the narrow area between the cervix and the corpus, and the endometrium in the cornual recesses at the ostia of the tubes remain intact). The new surface epithelium emanates from the flanks of stumps of glands in the basalis layer left standing after menstrual desquamation.17 Rapid re-epithelialization follows the proliferation of the cells in the basalis layer and the surface epithelium in the isthmic and tubal ostial endometrium. This epithelial repair is supported by underlying fibroblasts. The stromal fibroblast layer forms a compact mass over which the resurfacing epithelium can “migrate.” In addition, it is likely that the stromal layer contributes important autocrine and paracrine factors for growth and migration. Because hormone levels are at their nadir during this repair phase, the response may be due to injury rather than hormone mediated. However, the basalis layer is rich in its content of estrogen receptors. This “repair” is fast; by day 4 of the cycle, more than two-thirds of the cavity is covered with new epithelium.17 By day 5-6, the entire cavity is re-epithelialized, and stromal growth begins.
The Proliferative Phase
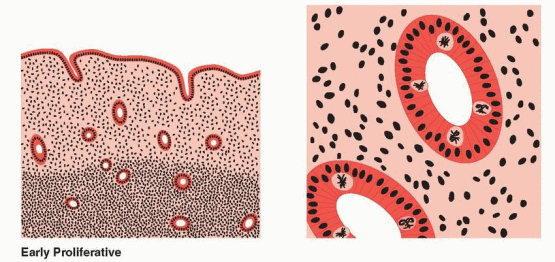
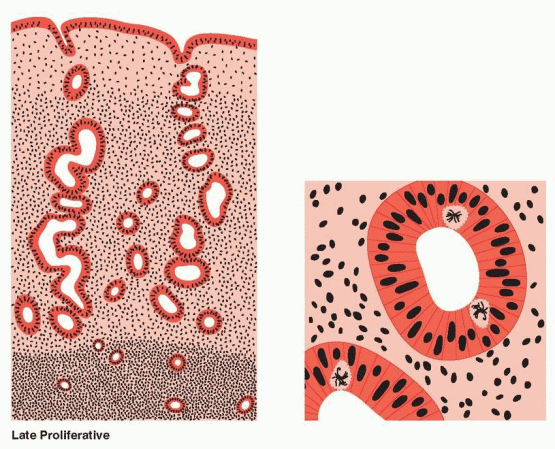
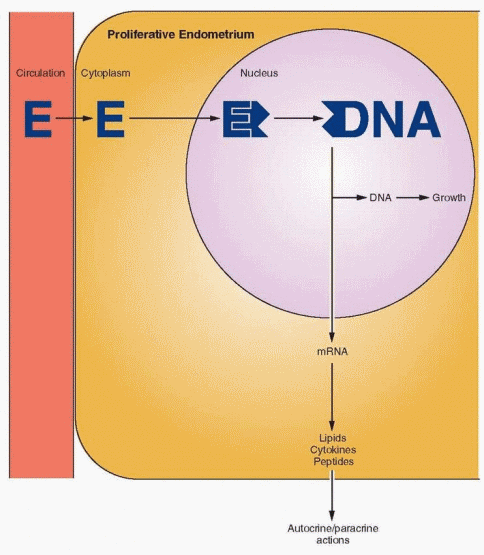
The proliferative phase is associated with ovarian follicle growth and increased estrogen secretion. Undoubtedly as a result of this steroidal action, reconstruction and growth of the endometrium are achieved. The glands are most notable in this response. At first they are narrow and tubular, lined by low columnar epithelium cells. Mitoses become prominent and pseudostratification is observed. As a result, the glandular epithelium extends peripherally and links one gland segment with its immediate neighbor. A continuous epithelial lining facing the endometrial cavity is formed. The stromal component evolves from its dense cellular menstrual condition through a brief period of edema to a final loose syncytial-like status. Coursing through the stroma, spiral vessels extend (unbranched and uncoiled in the early proliferative phase) to a point immediately below the epithelial binding membrane. Here they form a loose capillary network. All of the tissue components (glands, stromal cells, and endothelial cells) demonstrate proliferation, which peaks on days 8-10 of the cycle, reflecting rising estradiol levels in the circulation and maximal estrogen receptor concentration in the endometrium.18 This proliferation is marked by increased mitotic activity and increased nuclear DNA and cytoplasmic RNA synthesis, which is most intense in the functionalis layer in the upper two-thirds of the uterus, the usual site of blastocyst implantation.
During proliferation, the endometrium grows from approximately 0.5 mm to 3.5-5.0 mm in height of a singular layer. Restoration of tissue constituents has been achieved by estrogeninduced new growth as well as incorporation of ions, water, and amino acids. The stromal ground substance has re-expanded from its menstrual collapse. Although true tissue growth has occurred, a major element in achievement of endometrial height is “reinflation” of the stroma.
An important feature of this estrogen-dominant phase of endometrial growth is the increase in ciliated and microvillous cells. Ciliogenesis begins on days 7-8 of the cycle.17 This response to estrogen is exaggerated in hyperplastic endometrium that is the result of hyperestrogenism. The concentration of these ciliated cells around gland openings and the ciliary beat pattern influence the mobilization and distribution of endometrial secretions during the secretory phase. Cell surface microvilli, also a response to estradiol, are cytoplasmic extensions and serve to increase the active surface of cells.
At all times, a large number of cells derived from bone marrow are present in the endometrium. These include lymphocytes and macrophages, diffusely distributed in the stroma.
The Secretory Phase
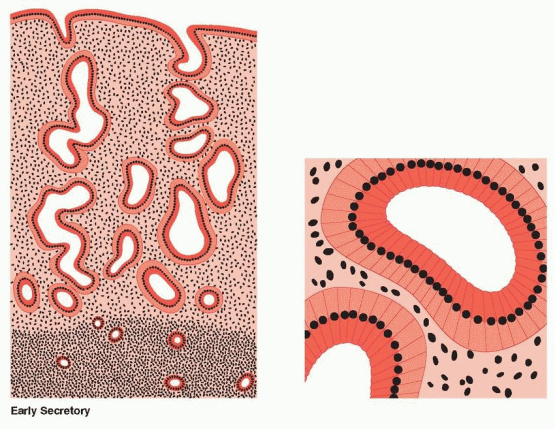
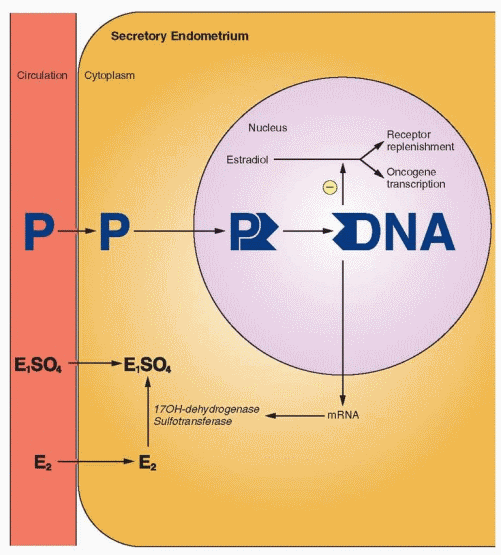
After ovulation, the endometrium now demonstrates a combined reaction to estrogen and progesterone activity. Most impressive is that total endometrial height is fixed at roughly its preovulatory extent (5-6 mm) despite continued availability of estrogen. Epithelial proliferation ceases 3 days after ovulation.19 This restraint or inhibition is believed to be induced by progesterone. This limitation of growth is associated with a decline in mitosis and DNA synthesis, significantly due to progesterone interference with estrogen receptor expression and progesterone stimulation of 17β-hydroxysteroid dehydrogenase and sulfotransferase, which convert estradiol to estrone sulfate (which is rapidly excreted from the cell).20,21 In addition, estrogen stimulates many oncogenes that probably mediate estrogeninduced growth. Progesterone antagonizes this action by suppressing the estrogen-mediated transcription of oncogene mRNA.22
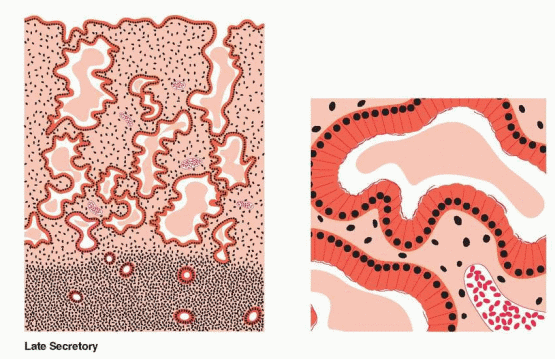
Individual components of the tissue continue to display growth, but confinement in a fixed structure leads to progressive tortuosity of glands and intensified coiling of the spiral vessels. The secretory events within the glandular cells, with progression of vacuoles from intracellular to intraluminal appearance, are well known and take place over a 7-day postovulatory interval. At the conclusion of these events, the glands appear exhausted, the tortuous lumina variably distended, and individual cell surfaces fragmented in a sawtooth appearance. Stroma is increasingly edematous, and spiral vessels are prominent and densely coiled.
The first histologic sign that ovulation has occurred is the appearance of subnuclear intracytoplasmic glycogen vacuoles in the glandular epithelium on cycle days 17-18. Giant mitochondria and the “nucleolar channel system” appear in the gland cells. The nucleolar channel system has a unique appearance due to progesterone, an infolding of the nuclear membranes. Individual components of the tissue continue to display growth, but confinement in a fixed structure leads to progressive tortuosity of glands and intensified coiling of the spiral vessels. These structural alterations are soon followed by active secretion of glycoproteins and peptides into the endometrial cavity. Transudation of plasma also contributes to the endometrial secretions. Important immunoglobulins are obtained from the circulation and delivered to the endometrial cavity by binding proteins produced by the epithelial cells. The peak secretory level is reached 7 days after the midcycle gonadotropin surge, coinciding with the time of blastocyst implantation.
The Implantation Phase
Significant changes occur within the endometrium from the 7th to the 13th day postovulation (days 21-27 of the cycle). At the onset of this period, the distended tortuous secretory glands have been most prominent with little intervening stroma. By 13 days postovulation, the endometrium has differentiated into three distinct zones. Something less than one-fourth of the tissue is the unchanged basalis fed by its straight vessels and surrounded by indifferent spindle-shaped stroma. The midportion of the endometrium (approximately 50% of the total) is the lace-like stratum spongiosum, composed of loose edematous stroma with tightly coiled but ubiquitous spiral vessels and exhausted dilated glandular ribbons. Overlying the spongiosum is the superficial layer of the endometrium (about 25% of the height) called the stratum compactum. Here the prominent histologic feature is the stromal cell, which has become large and polyhedral. In its cytoplasmic expansion one cell abuts the other, forming a compact, structurally sturdy layer. The necks of the glands traversing this segment are compressed and less prominent. The subepithelial capillaries and spiral vessels are engorged.
At the time of implantation, on days 21-22 of the cycle, the predominant morphologic feature is edema of the endometrial stroma. This change may be secondary to the estrogenand progesterone-mediated increase in prostaglandin and vascular endothelial growth factor (VEGF) production by the endometrium that cause an increase in capillary permeability. Receptors for the sex steroids are present in the muscular walls of the endometrial blood vessels, and the enzyme system for prostaglandin synthesis is present in both the muscular walls and the endothelium of the endometrial arterioles. Mitoses are first seen in endothelial cells on cycle day 22. Vascular proliferation leads to the coiling of the spiral vessels, a response to the sex steroids, the prostaglandins, and the autocrine and paracrine factors produced in response to estrogen and progesterone.
During the secretory phase, so-called K (Körnchenzellen) cells appear, reaching a peak concentration in the first trimester of pregnancy. These are granulocytes that have an immunoprotective role in implantation and placentation. They are located perivascularly and are believed to be derived from the blood. By day 26-27, the endometrial stroma is infiltrated by extravasated polymorphonuclear leukocytes. The majority of the leukocytes are killer cells and macrophages, believed to be involved in the process of endometrial breakdown and menstruation. The appearance and function of these cells are regulated by the complex array of peptides and cytokines in the endometrium in response to hormonal signaling.
The gene expression pattern in the endometrium throughout the menstrual cycle is being established, with a focus on the implantation window.23,24 and 25 As expected, microarray analyses reveal a changing pattern of gene expression that correlates with each hormonal and morphological stage in the endometrial menstrual cycle.26 Ultimately this will yield a comprehensive picture, with the gene signature of each event in the estrogen and progesterone regulation of the endometrium. The regulating growth factors, cytokines, and peptide hormones that are essential for implantation will be identified.
The stromal cells of the endometrium respond to hormonal signals, synthesize prostaglandins, and, when transformed into decidual cells, produce an impressive array of substances, some of which are prolactin, relaxin, renin, insulin-like growth factors (IGFs), and insulinlike growth factor binding proteins (IGFBPs). The endometrial stromal cells, the progenitors of decidual cells, were originally believed to be derived from the bone marrow (from cells invading the endometrium), but they are now considered to emanate from the primitive uterine mesenchymal stem cells.27
The decidualization process begins in the luteal phase under the influence of progesterone and mediated by autocrine and paracrine factors. On cycle days 22-23, predecidual cells can be identified, initially surrounding blood vessels, characterized by cytonuclear enlargement, increased mitotic activity, and the formation of a basement membrane. The decidua, derived from stromal cells, becomes an important structural and biochemical tissue of pregnancy. Decidual cells control the invasive nature of the trophoblast, and the products of the decidua play important autocrine and paracrine roles in fetal and maternal tissues.
Lockwood and his colleagues assign a key role to decidual cells in both the process of endometrial bleeding (menstruation) and the process of endometrial hemostasis (implantation and placentation).28,29 and 30 Implantation requires endometrial hemostasis and the maternal uterus requires resistance to invasion. Inhibition of endometrial hemorrhage can be attributed, to a significant degree, to appropriate changes in critical factors as a consequence of decidualization; e.g., lower plasminogen activator levels, reduced expression of the enzymes that degrade the stromal extracellular matrix (such as the metalloproteinases), and increased levels of plasminogen activator inhibitor-1. Withdrawal of estrogen and progesterone support, however, leads to changes in the opposite directions, consistent with endometrial breakdown.
The Phase of Endometrial Breakdown
Predecidual transformation has formed the “compacta” layer in the upper part of the functionalis layer by day 25 (3 days before menstruation). In the absence of fertilization, implantation, and the consequent lack of sustaining quantities of human chorionic gonadotropin from the trophoblast, the otherwise fixed lifespan of the corpus luteum is completed, and estrogen and progesterone levels wane.
The withdrawal of estrogen and progesterone initiates important endometrial events: vasomotor reactions, the process of apoptosis, tissue loss, and, finally, menstruation. The most prominent immediate effect of this hormone withdrawal is a modest shrinking of the tissue height and remarkable spiral arteriole vasomotor responses. The classic concept of the vascular sequence was constructed from direct observations of Rhesus endometrium transplanted to the anterior chamber of the eye.7,8 With shrinkage of height, blood flow within the spiral vessels diminished, venous drainage was decreased, and vasodilation ensued. Thereafter, the spiral arterioles underwent rhythmic vasoconstriction and relaxation. Each successive spasm was more prolonged and profound, leading eventually to endometrial blanching. Thus these reactions were proposed to lead to menstruation because of endometrial ischemia and stasis caused by vasoconstriction of the spiral arterioles. A new model of menstruation, as discussed in Chapter 15, emphasizes enzymatic autodigestion of the functional layer of the endometrium and its capillary plexus.
In the first half of the secretory phase, acid phosphatase and potent lytic enzymes are confined to lysosomes. Their release is inhibited by progesterone stabilization of the lysosomal membranes. With the waning of estrogen and progesterone levels, the lysosomal membranes are not maintained, and the enzymes are released into the cytoplasm of epithelial, stromal, and endothelial cells and eventually into the intercellular space. These active enzymes will digest their cellular constraints, leading to the release of prostaglandins, extravasation of red blood cells, tissue necrosis, and vascular thrombosis. This process is one of apoptosis, (programmed cell death, characterized by a specific morphologic pattern that involves cell shrinkage and chromatin condensation culminating in cell fragmentation) mediated by cytokines.31 An important step in this breakdown is the dissolution of cell-tocell adhesion by key proteins. Binding of endometrial epithelial cells utilizes transmembrane proteins, cadherins, that link intercellularly with each other and intracellularly with catenins that are bound to actin filaments.32
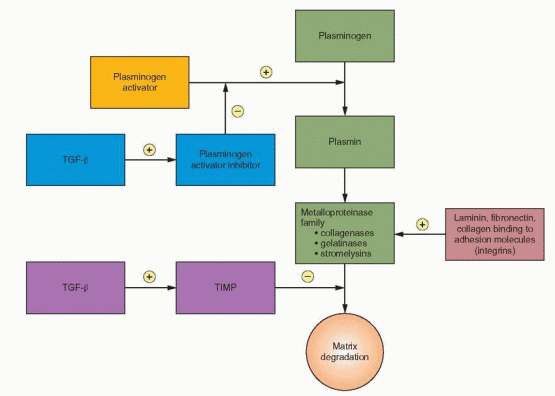
Endometrial tissue breakdown also involves a family of enzymes, matrix metalloproteinases, that degrade components (including collagens, gelatins, fibronectin, and laminin) of the extracellular matrix and basement membrane.33,34 The metalloproteinases include collagenases that degrade interstitial and basement membrane collagens; gelatinases that further degrade collagens; and stromelysins that degrade fibronectin, laminin, and glycoproteins. The expression of metalloproteinases in human endometrial stromal cells follows a pattern correlated with the menstrual cycle, indicating a sex steroid response as part of the growth and remodeling of the endometrium with a marked increase in late secretory and early menstrual endometrium.35 Progesterone withdrawal from endometrial cells increases VEGF production and induces matrix metalloproteinase secretion, probably from both endometrial stromal cells and leukocytes, which is followed by the irreversible breakdown of cellular membranes and the dissolution of extracellular matrix.36,37 and 38 Appropriately, this enzyme expression increases in the decidualized endometrium of the late secretory phase, during the time of declining progesterone levels. With the continuing progesterone secretion of early pregnancy, the decidua is maintained and metalloproteinase expression is suppressed, in a mechanism mediated by transforming growth factor-beta (TGF-β).39 In a nonpregnant cycle, metalloproteinase expression is suppressed after menses, presumably by increasing estrogen levels.
Metalloproteinase activity is restrained by specific tissue inhibitors designated as TIMP.40 The balance of metalloproteinase and TIMP activity is an important event in successful implantation. Thus, progesterone withdrawal can lead to endometrial breakdown through a mechanism that is independent of vascular events (specifically ischemia), a mechanism that involves cytokines.31 During bleeding, both normal and abnormal, there is evidence indicating that specific genes are activated in the endometrium; one such gene has the structural features of the TGF-β family.41
There is considerable evidence to support a major role for a cytokine, tumor necrosis factor-a (TNF-a), in menstruation.31 TNF-a is a transmembrane protein whose receptor belongs to the nerve growth factor/TNF family for inducing apoptotic signals. The key change is an increase in secretion because TNF-a secretion by endometrial cells reaches a peak at menstruation, but there is no cycle change in receptor content. TNF-a inhibits endometrial proliferation and induces apoptosis; this cytokine causes a loss of adhesion proteins (the cadherin-catenin-actin complex) and induces cell-to-cell dissolution. In addition to endometrial cells, TNF-a also causes damage to vascular endothelium.
Progesterone withdrawal is also associated with an increase in vascular endothelial growth factor receptor concentrations in the stromal cells of the layers of endometrium destined to be sloughed.42 Although the vascular endothelial growth factor system is usually involved with angiogenesis, in this case these factors are involved in the preparation for menstrual bleeding, perhaps influencing the expression of matrix metalloproteinases (MMPs). Endometrial genes without classic steroid response elements can respond to the sex steroids by utilizing a family of proteins (the Sp family) that mediate steroid activity at the level of transcription (acting in a fashion similar to the steroid receptors). These proteins, induced by progesterone in stromal (decidual) and epithelial cells, can activate tissue factor, plasminogen activator inhibitor-1, IGF binding protein-1, uteroglobin, and uteroferrin. Tissue factor is involved in the clotting mechanism to sustain hemostasis. Uteroglobin is a small protein expressed in endometrial epithelial cells.43 The physiologic function of uteroglobin is uncertain. Uteroglobin, with high affinity, binds progestins and may play a role in immunosuppression. Uteroglobin gene expression is stimulated by estrogen, and this response is enhanced by progesterone. Human endometrium can secrete β-endorphin, yet another candidate for involvement in endometrial immunologic events, and its release is inhibited by both estrogens and glucocorticoids.44
Eventually, considerable leakage occurs as a result of diapedesis, and finally, interstitial hemorrhage occurs due to breaks in superficial arterioles and capillaries. White cells migrate through capillary walls, at first remaining adjacent to vessels but then extending
throughout the stroma. The leukocytes add important regulatory cytokines, chemokines, and enzymes that are involved in the degradation of the extracellular matrix. During arteriolar vasomotor changes, red blood cells escape into the interstitial space. Thrombinplatelet plugs also appear in superficial vessels. The prostaglandin content (PGF2a and PGE2) in the secretory endometrium reaches its highest levels at the time of menstruation. The vasoconstriction and myometrial contractions associated with the menstrual events are believed to be significantly mediated by prostaglandins from perivascular cells and the potent vasoconstrictor endothelin-1, derived from stromal decidual cells.
As ischemia and weakening progress, the continuous binding membrane is fragmented, and intercellular blood is extruded into the endometrial cavity. New thrombin-platelet plugs form intravascularly upstream at the shedding surface, limiting blood loss. Increased blood loss is a consequence of reduced platelet numbers and inadequate hemostatic plug formation. Menstrual bleeding is influenced by activation of clotting and fibrinolysis. Fibrinolysis is principally the consequence of the potent enzyme plasmin, formed from its inactive precursor plasminogen. Endometrial stromal cell tissue factor (TF) and plasminogen activators and inhibitors are involved in achieving a balance in this process. TF stimulates coagulation, initially binding to factor VII. TF and plasminogen activator inhibitor-1 (PAI-1) expression accompanies decidualization, and the levels of these factors may govern the amount of bleeding.30,45 PAI-1, in particular, exerts an important restraining action on fibrinolysis and proteolytic activity.46 Blood loss is also controlled by constriction of the spiral arteries, mediated by the perivascular cells, myofibroblasts that surround the spiral arteries.47 These cells respond to progesterone withdrawal by expressing prostaglandins, cytokines, and MMPs, causing not only cycling vasoconstriction and vasodilation but also modulating leukocyte entry (an important additional source of metalloproteinases) into the endometrium. Disordered growth and function of the perivascular cells are likely contributing factors in menstrual bleeding problems.
High Progesterone Levels
Progesterone Withdrawal
↓
↓
Perivascular Growth and Decidualization
Prostaglandin, Cytokine, and VEGF Expression
↓
↓
Suppression of Prostaglandin, Cytokine, and MMP Expression
Vasoconstriction, Vasodilation, Leukocyte Infiltration, and Increase in MMPs
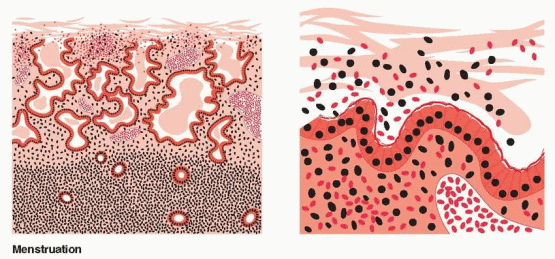
With progressive enzymatic degradation of the endometrium, the subsurface capillary and venous vascular system is disrupted, causing hemorrhage and escape of blood into the endometrial cavity. Additional ischemic breakdown ensues with necrosis of cells and defects in vessels adding to the menstrual effluvium. Degeneration extends to the deepest extent of the functional layer where rupture of the basal arterioles contributes to bleeding. A natural cleavage point exists between basalis and spongiosum, and, once breached, the loose, vascular, edematous stroma of the spongiosum desquamates and collapses. The process is initiated in the fundus and inexorably extends throughout the uterus. In the end, the typical deflated, shallow, dense, menstrual endometrium results. Within 13 hours, the endometrial height shrinks from 4 to 1.25 mm.13 Menstrual flow stops as a result of the combined effects of prolonged vasoconstriction of the radial arteries and the spiral arteries in the basalis, tissue collapse, vascular stasis, and estrogeninduced “healing.” In contrast to postpartum bleeding, myometrial contractions are not important for control of menstrual bleeding. Thrombin generation in the basal endometrium in response to extravasation of blood is essential for hemostasis. Thrombin promotes the generation of fibrin, the activation of platelets and clotting cofactors, and angiogenesis.
The basalis endometrium remains during menses, and repair takes place from this layer. This endometrium is protected from the lytic enzymes in the menstrual fluid by a mucinous layer of carbohydrate products that are discharged from the glandular and stromal cells.48
Normal Menses
Approximately 50% of the menstrual detritus is expelled in the first 24 hours of menstrual flow. The menstrual fluid is composed of the autolysed functionalis, inflammatory exudate, red blood cells, and proteolytic enzymes (at least one of which, plasmin, lyses fibrin clots as they form). The high fibrinolytic activity advances emptying of the uterus by liquefaction of tissue and fibrin. If the rate of flow is great, clotting can and does occur.
Most women (90%) have menstrual cycles with an interval of 24 to 35 days (Chapter 6).49,50 Menarche is followed by approximately 5-7 years of increasing regularity as cycles shorten to reach the usual reproductive-age pattern. In the 40s, cycles begin to lengthen again. The usual duration of flow is 4-6 days, but many women flow as little as 2 days and as much as 8 days. The normal volume of menstrual blood loss is 30 mL; greater than 80 mL is abnormal. Normal and abnormal characteristics and definitions of menstrual flow are discussed in detail in Chapter 15.
A Teleologic Theory of Endometrial-Menstrual Events
Menstruation is a very recent phenomenon in the evolutionary time line. It occurs in very few species, even among viviparous animals. An unabashedly teleologic view of menstrual events was offered many years ago by Rock et al.51 The basic premise of this thesis is that every endometrial cycle has, as its only goal, nourishing support of an early embryo. Failure to accomplish this objective is followed by orderly elimination of unutilized tissue and prompt renewal to achieve a more successful cycle.
The ovum must be fertilized within 12-24 hours of ovulation. Over the next 4 days, it remains unattached within the tubal lumen utilizing tubal fluids and residual cumulus cells to sustain nutrition and energy for early cellular cleavage. After this stay, the solid ball of cells (morula), which is the embryo, leaves the tube and enters the uterine cavity. Here the embryo undergoes another 2-3 days of unattached but active existence. Fortunately, by this time endometrial gland secretions have filled the cavity and they bathe the embryo in nutrients. This is the first of many neatly synchronized events that mark the conceptus-endometrial relationship. By 6 days after ovulation, the embryo (now a blastocyst) is ready to attach and implant. At this time, it finds an endometrial lining of sufficient depth, vascularity, and nutritional richness to sustain the important events of early placentation to follow. Just below the epithelial lining, a rich capillary plexus has been formed and is available for creation of the trophoblast-maternal blood interface. Later, the surrounding zona compactum, occupying more and more of the endometrium, will provide a sturdy splint to retain endometrial architecture despite the invasive inroads of the burgeoning trophoblast.
Failure of the appearance of human chorionic gonadotropin, despite otherwise appropriate tissue reactions, leads to the vasomotor changes associated with estrogen-progesterone withdrawal and menstrual desquamation. However, not all the tissue is lost, and, in any event, a residual basalis is always available, making resumption of growth with estrogen a relatively rapid process. Indeed, even as menses persists, early regeneration can be seen. As soon as follicle maturation occurs (in as short a time as 10 days), the endometrium is ready once again to perform its reproductive function.
The Uterus Is an Endocrine Organ
The uterus is dynamic. It not only responds and changes in a sensitive fashion to classic hormonal signals (the endocrine events of the menstrual cycle) but it is also composed of complex tissues, with important autocrine and paracrine functions that serve not only the uterus but also the contiguous tissues of the fetoplacental unit during pregnancy. The most dynamic component of the uterus is the endometrium.
Endometrial Products
The endometrium secretes many substances, the functions of which (and their interrelationships) represent a major investigative challenge.52 In addition to producing a nourishing, supportive environment for the early embryo, the endometrium plays an important role in suppressing the immune response within the pregnant uterus. The mechanisms controlling the immune response in decidual cells are not understood, but hormonal influence is undoubtedly important.
The presence of the cytokine family, involved in inflammation and immune responses, is not surprising in a tissue that undergoes cyclic degeneration. The interleukins stimulate the production of prostaglandins as well as other cytokines.53 Colony-stimulating factor-1 is a cytokine that influences cellular proliferation and the presence of macrophages. Interferon-γ is produced by activated T lymphocytes and inhibits endometrial epithelial proliferation. Leukemia-inhibiting factor (LIF) is expressed in response to a variety of other cytokines and growth factors. Like the interleukins, LIF is most abundant during the progesteronedominated secretory phase and early decidua and may have a role in embryo implantation.54,55 Tumor necrosis factor-a (TNF-a) gene expression is present in endometrium, and its activity is increased during the proliferative phase, decreased early in the secretory phase, and increased again in the midsecretory phase.56 TNF-a exerts multiple influences on cellular growth.
A Partial List of Endometrial Regulating Molecules
Lipids
Cytokines
Peptides
Prostaglandins
Interleukin-1α
Prolactin
Thromboxanes
Interleukin-1β
Relaxin
Leukotrienes
Interleukin-6
Prorenin and Renin
Interferon-γ
Endorphin
Colony-stimulating factor-1
Endothelin-1
Tumor necrosis factor-α
Corticotropin-releasing hormone
Leukemia-inhibiting factor
Fibronectin
Uteroglobin
Lipocortin-1
Parathyroid hormone-like protein
Integrins
Epidermal growth factor family
EGF
Heparin-binding EGF
TGF-α
Insulin-like growth factor family
IGF-I
IGF-II
IGFBPs 1-6
Transforming growth factor-β family
Activins
Inhibins
Follistatin
Platelet-derived growth factor
Fibroblast growth factor
Vascular endothelial growth factor
Gonadotropin-releasing hormone (GnRH)
Growth factors are peptides that bind to specific cell membrane receptors and initiate intracellular signaling pathways. Because the growth factors are potent mitogens, it is also not surprising that the follicular phase of the cycle, associated with proliferative activity of the endometrium, is marked by dramatic alterations in growth factors. Estrogen stimulates gene expression for epidermal growth factor (EGF) (and its receptor) and insulin-like growth factor (IGF) production. In turn, EGF elicits estrogen-like actions by interacting with the estrogen receptor mechanism.57 EGF, a potent mitogen, is present in endometrial stromal and epithelial cells during the follicular phase of the cycle and in the stromal cells during the luteal phase.58 Transforming growth factor-a (TGF-a) and EGF work through the same receptor and are important mediators of estrogen-induced growth of the endometrium. TGF-a levels peak at midcycle, in contrast to EGF levels, which are relatively stable and noncyclic.59,60 and 61 Platelet-derived growth factor is a potent mitogen localized to stromal cells.
The insulin-like growth factors promote cellular mitosis and differentiation. They are expressed in a pattern controlled by estrogen and progesterone. IGF-I is predominant in proliferative and early secretory endometrium, and IGF-II appears in the mid to late secretory phase and persists in early pregnancy decidua.62 Endometrial IGF-I expression is correlated with the circulating estrogen levels during the menstrual cycle.63 This suggests that IGF-I synthesis is regulated by estrogen and mediates estrogen-induced growth of the endometrium, and IGF-II is involved in differentiation in response to progesterone. Evidence in the monkey indicates that IGF-I is the primary regulator of myometrial growth in response to estrogen as well as to estrogen plus progesterone.64
As elsewhere in the body, the myometrial IGF activity is modulated by the IGF binding proteins, which respond to the sex steroids in a differential manner; IGFBP-2 parallels IGF-I response, whereas IGFBP-3 is decreased in muscle but increased in vascular endothelium by estrogen.65 IGFBP-4 and IGFBP-5 respond to estrogen but are unaffected by the addition of progesterone. IGFBP-1, as discussed later, is a major product of decidualized endometrium.
Gonadotropin-releasing hormone (GnRH) is present in endometrium and in increased amounts in secretory endometrium and decidua.66 In human decidual cells, GnRH increases the expression of matrix metalloproteinases, suggesting a role for GnRH in the regulation of the enzymes involved in implantation.67 Like all of these molecules, GnRH is involved in signaling pathways associated with cell proliferation and breakdown, interacting with adhesion factors such as integrins, enzymes, and angiogenic substances.68
Human myometrial smooth muscle and endometrial stromal cells express mRNA for parathyroid hormone-like protein, the function of which is unknown.69 Transforming growth factor-β (TGF-β) stimulates the production of the parathyroid hormone-like protein. TGF-β production is greatest in the secretory phase and may inhibit cellular proliferation by increasing IGFBP-3 synthesis.
Prostaglandins are produced by both epithelial and stromal cells, and the prostaglandin content in the endometrium reaches a peak level in late secretory endometrium. The predominant prostaglandin produced by endometrium is prostaglandin F2a, a potent stimulus for myometrial contractions.70 Endometrial prostaglandin production decreases dramatically after implantation, suggesting the presence of an active mechanism for suppression.71 The production of prostaglandins requires estrogen support, but the increased production by secretory endometrium indicates progesterone enhancement, and acute withdrawal of progesterone promotes a further increase.70 Endometrial stromal cells produce prostacyclin and thromboxane in response to estrogen, a response that can be blocked by progestins.72 The myometrium principally produces prostacyclin, utilizing precursors derived from the endometrium. However, receptors for all members of the prostaglandin family are present on human myometrial cells, and contraction of the myometrium is a major consequence of prostaglandin F2a.73
Thromboxane is synthesized by uterine tissues. Gene expression for the thromboxane synthase and for the thromboxane receptor can be identified in endometrial glands, stromal cells, myometrial smooth muscle, and uterine blood vessels.74 Thromboxane A2 is a potent vasoconstrictor and stimulator of smooth muscle cells. Because of its rapid metabolism, it is limited to autocrine and paracrine activity.
Women with excessive menstrual bleeding have alterations in the normal rates of prostaglandin production. For this reason, effective reductions in menstrual blood loss can be achieved with treatment utilizing one of the nonsteroidal anti-inflammatory agents that inhibit prostaglandin synthesis. These agents are also effective treatment for prostaglandinmediated dysmenorrhea.
Fibronectin and laminin are extracellular matrix substances that are secreted by stromal cells of the endometrium in response to progesterone.75 These proteins are important adhesion molecules during implantation. Integrins are a family of glycoproteins that function as receptors for proteins such as collagen, fibronectin, and laminin. The integrins are highly expressed in endometrium and are important for cell-to-cell and cell-to-matrix interactions.76 The expression of integrins is regulated by cytokines and growth factors, not estrogen and progesterone.77
Endothelins are potent vasoconstrictors produced in the vascular endothelial cells. The vasoconstrictor activity of endothelin-1, present in the endometrium, is balanced by the fact that it promotes the synthesis of the vasodilators nitric oxide and prostacyclin. Endothelin-1 is synthesized in endometrial stromal cells and the glandular epithelium, stimulated by both TGF-β and interleukin-1a.78 Endothelin-1 is at least one agent responsible for the vasoconstriction that shuts off menstrual bleeding. It is also a potent stimulator of myometrial contractions and can contribute to dysmenorrhea. Finally, endothelin-1 is a mitogen and can promote the healing re-epithelialization of the endometrium. Human decidual cells also synthesize and secrete endothelin-1, from where it may be transported into the amniotic fluid.79
Angiogenesis, the formation of new blood vessels, is an essential process in tissue growth and development. Angiogenesis is necessary for tumor growth, and, in normal tissues, it is usually kept in check by regulating factors. The female reproductive tissues (specifically ovarian follicles, the trophoblast, and the endometrium), however, must experience periodic and rapid growth and regression. In these tissues, angiogenesis is part of normal events. The endometrium is a major source for angiogenic factors during the menstrual cycle and during pregnancy.80 Vascular endothelial growth factors (VEGFs), a collection of specific mitogens for endothelial cells, are abundantly expressed in human endometrium, reaching a peak that correlates with the maximal angiogenesis reached during the secretory phase.81,82 The VEGF family contains six growth factors and utilizes three different receptors. During the proliferative phase, estrogen stimulates VEGF synthesis. VEGF expression is also stimulated by hypoxia, specifically the hypoxia associated with endometrial breakdown, and the new blood vessel growth as well as the re-epithelialization of the endometrium in the new proliferative phase are dependent on these growth factors in response to estrogen.83,84
Angiogenesis is also influenced by many other growth factors and other substances such as fibronectin and prostaglandins. The fibroblast growth factor family, in particular, is highly mitogenic for endothelial cells and endometrial stromal cells. Angiopoietins sustain the endometrium by preventing apoptosis and stabilizing blood vessels. The endometrium also produces inhibitory proteins, and the final growth of blood vessels reflects the balance between the inhibitory and stimulatory factors.
In all types of endometrial and myometrial cells, estrogen receptor expression reaches a maximum in the late proligerative phase.85,86 The concentration is greatest in the glandular epithelium. During the early secretory phase, estrogen receptor expression declines, followed by an increase in the mid and late secretory phases. These changes reflect the cyclic changes in estradiol (which increases estrogen receptor expression) and progesterone (which decreases estrogen receptor expression). Although estrogen receptor-beta is present in human endometrium, it is less prominent than estrogen receptor-alpha and exhibits less change during the cycle, except when it becomes the predominant estrogen receptor in the endometrial vasculature in the late secretory period.87 Estrogen stimulation of proliferation is largely, if not totally, mediated by estrogen receptor-alpha.
Progesterone receptor expression in endometrial glandular epithelium reaches a maximum in the late proliferative and early secretory phases (reflecting induction of progesterone receptor by estrogen) and then declines to nearly undetectable levels by the midpoint of the secretory phase. Stromal cells in the endometrium show only minor fluctuations in progesterone receptors during the menstrual cycle. Decidualizing stromal cells exhibit strong progesterone receptor expression, although progesterone receptors are absent from decidual epithelial cells. Smooth muscle cells of the uterus demonstrate strong progesterone receptor expression throughout the menstrual cycle. Many of the events in uterine growth and function are regulated by the interplay between estrogen and progesterone. In general, progesterone antagonizes estrogen stimulation of proliferation and metabolism. This antagonism can be explained by the effects of progestins on the estrogen receptor (a decrease in levels), on the enzymes that lead to excretion of estrogen from cells, and by progesterone suppression of estrogen-mediated transcription of oncogenes.
Androgen receptor is present in endometrium at all stages of the menstrual cycle, in postmenopausal endometrium, and in the decidua of pregnancy.88 Surprisingly, the androgen receptor concentration is constant throughout the cycle. Androgens suppress the proliferative effects of estrogen on the endometrium, and experimental evidence suggests that the suppressive effects on the endometrium induced by antiprogestational agents are mediated by the androgen receptor.89
The complexity of the endometrium can be appreciated by viewing the results of complementary DNA microarray studies. In one effort directed just to the endometrial breakdown associated with menstruation, 571 transcripts were identified that were involved in 131 biochemical pathways, including thyroid hormone synthesis and metabolism!90 Gene expression studies are just beginning to profile the patterns associated with specific hormones and pharmacologic agents.91
The Decidua
The decidua is the specialized endometrium of pregnancy. The biochemical dialogue between the fetoplacental unit and the mother must pass back and forth through the decidua. The classic view of the decidua conformed to its designation as a thin line in anatomic diagrams, a minor, inactive structural component. We now know that the decidua is a vigorous, active tissue.
Decidual cells are derived from the stromal cells of the endometrium, under the stimulation of progesterone. This transformation is regulated by members of the transforming growth factor beta family, including activin A.92,93 In addition, ghrelin acting via the growth hormone receptor is involved in this process.94
Decidual cells appear during the secretory phase and continue to proliferate during early pregnancy, eventually lining the entire uterus including the implantation site. The decidual cell is characterized by the accumulation of glycogen and lipid droplets and the new expression of a host of substances, including prolactin, relaxin, renin, insulin-like growth factors (IGFs), and insulin-like growth factor binding proteins (IGFBPs). There is no evidence that these proteins are secreted into the circulation; therefore, they serve as autocrine and paracrine agents.95,96
Riddick was the first to detect prolactin in the decidualizing endometrium of the late luteal phase.97 The amino acid sequence and the chemical and biologic properties of decidual prolactin are identical to those of pituitary prolactin. Decidual prolactin synthesis and release are controlled by the placenta, fetal membranes, and decidual factors. Dopamine, bromocriptine, and thyrotropin-releasing hormone (TRH), in contrast to their action in the pituitary, have no effect on decidual synthesis and release of prolactin. A protein named decidual prolactin-releasing factor has been purified from the placenta, and an inhibiting protein, which blocks the stimulatory activity of the releasing factor, has been purified from decidua.96 IGF-1, relaxin, and insulin all stimulate decidual prolactin synthesis and release, each through its own receptor. The same decidual cells produce both prolactin and relaxin. Prolactin exerts an overall inhibitory effect on the process of decidualization, perhaps an autocrine mechanism to limit the extent of decidualization.98
Lipocortin-1 is a calcium- and phospholipid-binding protein, present in the placenta and decidua, that inhibits phospholipase A2 and responds to glucocorticoids. Lipocortin-1 inhibits decidual prolactin release but in a mechanism independent of phospholipase action and independent of glucocorticoids. The prostaglandin system is not involved in decidual prolactin production, and corticosteroids do not affect decidual prolactin release.99
There is good reason to believe that amniotic fluid prolactin is derived from the decidua. In vitro experiments indicate that the passage of prolactin across the fetal membranes is in the direction of the amniotic cavity. The amniotic fluid concentration correlates with the decidual content, not with maternal circulating levels. Amniotic fluid prolactin reaches peak levels in the first half of gestation (about 4,000 ng/mL) when maternal plasma levels are approximately 50 ng/mL and fetal levels about 10 ng/mL. Maternal circulating prolactin reaches maximal levels near term. Finally, amniotic fluid prolactin is unaffected by bromocriptine treatment (which reduces both fetal and maternal circulating levels to baseline levels).
It is believed that decidual prolactin regulates amniotic fluid volume and electrolyte concentrations. Prolactin regulates water and ion transport in lower animals, and prolactin binds to amniotic membranes. Disorders in human pregnancy associated with abnormal amniotic fluid volumes may be explained by this mechanism, especially idiopathic polyhydramnios, which is associated with a decrease in the number of prolactin receptors in the membranes. Prolactin may be involved in the regulation of surfactant synthesis in the fetus, and prolactin may inhibit uterine muscle contractility. Prolactin suppresses the immune response and helps to prevent immunologic rejection of the conceptus. Prolactin can also function as an autocrine and paracrine growth factor in the uterus.100
Only gold members can continue reading. Log In or Register to continue