Chapter 3
Uteroplacental Blood Flow
Warwick D. Ngan Kee BHB, MBChB, MD, FANZCA, FHKCA, FHKAM
Chapter Outline
CHANGES AND FUNCTION DURING PREGNANCY
MECHANISMS OF VASCULAR CHANGES AND REGULATION
METHODS OF MEASUREMENT OF UTEROPLACENTAL BLOOD FLOW
Uteroplacental blood flow is responsible for the delivery of oxygen and nutrients to the fetus. A normal uteroplacental circulation is essential for healthy fetal growth and development. Acute reduction in uteroplacental blood flow may rapidly threaten fetal viability. Chronic reduction in uteroplacental blood flow, as may occur from abnormal development of the placental vasculature, leads to gestational pathologic processes such as preeclampsia and fetal growth restriction (also known as intrauterine growth restriction) and may even predispose the fetus to developing cardiovascular disease during subsequent adulthood.1 The uteroplacental circulation undergoes circadian changes2 and may be affected by parturition, disease, and anesthetic techniques and drugs. An understanding of the regulation of uteroplacental circulation is an important foundation for the safe provision of obstetric anesthesia and assists in the management of many pregnancy-related diseases. Research in this area is active but complicated by ethical considerations. Much of the available knowledge comes from studies in animals, particularly sheep but also nonhuman primates and other species. It is important to consider possible interspecies differences and to critically examine the methodology and context of animal research when extrapolating findings into recommendations for clinical care.
Anatomy and Structure
The blood supply to the uterus is derived mainly from the uterine arteries (Figure 3-1) with a smaller, variable contribution from the ovarian arteries. Although the pelvic vasculature shows anatomic variation,3 the uterine artery arises bilaterally from the anterior division of the internal iliac (hypogastric) artery, whereas the ovarian artery arises from the anterolateral abdominal aorta below the renal arteries. The uterine artery passes medially to the side of the uterus, where it supplies branches to the cervix and vagina and ascends between the two layers of the broad ligament, yielding arcuate arteries that supply the body of the uterus to the junction with the fallopian tubes. During pregnancy, flow may differ between the right and left uterine arteries; Konje et al.4 estimated that vessel diameter was approximately 11% greater and blood flow was approximately 18% greater in the uterine artery on the same side as the placenta compared with the contralateral artery. Anastomoses are formed with the contralateral uterine artery, the vaginal arteries, and the ovarian arteries. The arcuate arteries give rise to small branches that supply the myometrium and large radial arteries that branch deeply and enter the endometrium to form the convoluted spiral arteries. During gestation, trophoblastic invasion of the spiral arteries results in loss of smooth muscle and loss of contractile ability, leading to vasodilation with decreased resistance and increased blood flow. Abnormal or inadequate trophoblastic invasion is integral to the pathophysiology of preeclampsia (see Chapter 36).5
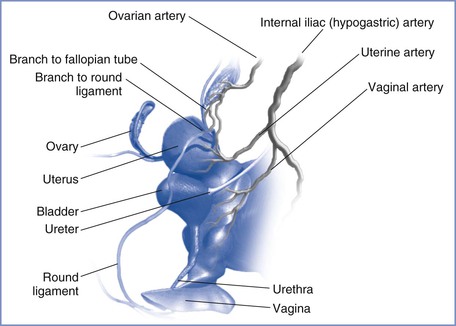
FIGURE 3-1 Arterial supply to the female reproductive tract. (Drawing by Naveen Nathan, MD, Northwestern University Feinberg School of Medicine, Chicago, IL.)
From the spiral arteries, oxygenated maternal blood enters the intervillous space in fountain-like jets. Blood traveling toward the chorionic plate bathes the villi, permitting the exchange of oxygen, nutrients, and wastes between maternal and fetal blood. Maternal blood then returns to the basal plate and drains into multiple collecting veins. Venous drainage of the uterus occurs via the uterine veins to the internal iliac veins and also via the ovarian veins (utero-ovarian plexus) to the inferior vena cava on the right and the renal vein on the left.6 The uterine artery and other branches of the anterior division of the internal iliac artery, as well as the ovarian artery, may be targeted during angiographic embolization procedures for treatment of obstetric and gynecologic hemorrhage3 and for the treatment of uterine fibroids.7
Changes and Function during Pregnancy
Pregnancy-Induced Changes
Uterine blood flow increases dramatically during pregnancy, rising from 50 to 100 mL/min before pregnancy to 700 to 900 mL/min at term, depending on the method of measurement (Figure 3-2). Studies in sheep have shown that increases in uterine blood flow can be divided into three phases.8 An initial phase, most likely controlled by the ovarian hormones estrogen and progesterone, occurs before and during implantation and early placentation. A second phase results from the growth and remodeling of the uteroplacental vasculature to support further placental development. A third and final phase results from progressive uterine artery vasodilation to meet the markedly increased nutrient requirements of the rapidly growing fetus. When expressed in terms of uterine weight, however, uterine flow per gram of tissue is particularly high in early gestation, and this ratio decreases as pregnancy progresses.8 In comparison, umbilical blood flow, expressed as a function of fetal weight, is relatively constant throughout most of pregnancy and is estimated to be 110 to 120 mL/min/kg.9 Uterine blood flow is increased in twin pregnancy, but the flow per unit of estimated fetal weight is similar to that in a singleton pregnancy.10 The progressive increase in uteroplacental blood flow during pregnancy is matched by a concurrent increase in blood flow on the fetal side (fetoplacental blood flow). However, despite suggestions of the possibility of intrinsic flow matching, it is believed that these circulations are independently regulated.11
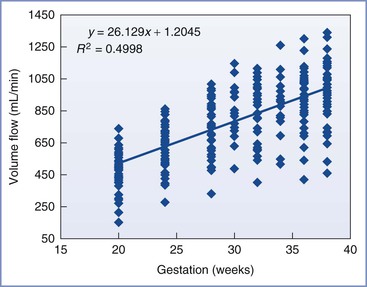
Distribution of Blood Flow
Uterine blood flow at term represents a greater proportion of cardiac output (approximately 12%) than in early pregnancy (approximately 3.5%).12 Regional distribution of blood flow within the pelvis also changes during gestation. Palmer et al.13 observed that increases in common iliac artery blood flow during pregnancy were associated with corresponding increases in uterine artery blood flow but also with decreases in external iliac artery blood flow. This pattern effectively constitutes a “steal” phenomenon, in which blood flow in the pelvis is preferentially redistributed toward the uterus (Figure 3-3).
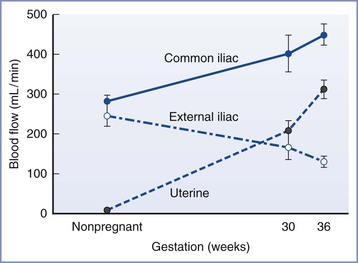
FIGURE 3-3 Redistribution of blood flow in pelvic blood vessels during pregnancy determined unilaterally by Doppler ultrasonography. Blood flow increased in the common iliac and uterine arteries but decreased in the external iliac artery, indicating that redistribution of flow favors uterine perfusion. Data are mean ± SEM. (Adapted from Palmer SK, Zamudio S, Coffin C, et al. Quantitative estimation of human uterine artery blood flow and pelvic blood flow redistribution in pregnancy. Obstet Gynecol 1992; 80:1000-6.)
Primate studies have shown that 80% to 90% of total uterine blood flow perfuses the placenta at term, with the remainder supplying the myometrium and nonplacental endometrium.14 The placental and nonplacental vasculatures are anatomically and functionally distinct, and regulation of perfusion through these vascular beds differs.14 Therefore, it is important to differentiate studies that measure total uteroplacental blood flow versus placental blood flow.
Functional Classification
Placental vascular function varies among species. The human multivillous model is commonly believed to function as a “venous equilibrator,” in which oxygen tension in the umbilical vein approximates that in the uterine veins. In contrast, the placenta in some species (e.g., rodents) functions as a countercurrent exchanger. The more efficient function of the latter is reflected by the higher fetoplacental weight ratio in rodents (20 : 1) than in humans (6 : 1).15
Autoregulation
Studies of pressure-flow relationships suggest that the nonpregnant uterine circulation exhibits autoregulation, alternately vasoconstricting or vasodilating in response to a number of different stimuli.16 In contrast, the pregnant uterine circulation is complicated by the properties of both the placental and nonplacental circulations. Animal studies have demonstrated that the uteroplacental circulation is a widely dilated, low-resistance system with perfusion that is largely pressure dependent.17,18 However, a study in pregnant rabbits found that uteroplacental blood flow was relatively constant over a wide range of perfusion pressures.19 During hemorrhage in pregnant rats, uterine vascular resistance increased as systemic blood pressure and uterine blood flow decreased, thereby demonstrating an absence of autoregulation. Moreover, although the uteroplacental circulation is often considered to be maximally vasodilated with little or no ability for autoregulation,17 further vasodilation has been observed in response to systemically administered estrogen, prostacyclin, bradykinin, and acetylcholine.20–22 These discrepancies may be explained by changes in the nonplacental uterine vasculature, which accounts for a small fraction of total uteroplacental blood flow but appears to have similar autoregulatory responses during pregnant and nonpregnant states; this feature contrasts with the limited autoregulatory ability of the placental circulation.23 Laird et al.18 found that reducing arterial pressure by 22% with an inflatable aortic occluder in pregnant rabbits produced a reduction in total uteroplacental and placental blood flow but no significant change in myoendometrial blood flow. Clinically, limited autoregulation means that placental blood flow may diminish with reductions in maternal blood pressure (e.g., during neuraxial anesthesia).
Margin of Safety
Studies in animals have demonstrated that, in normal physiologic conditions, uterine blood flow exceeds the minimum required to satisfy fetal oxygen demand.24 Although this feature confers a margin of safety that protects the fetus from fluctuations in uterine blood flow,25 decreases in fetal PO2 and progressive metabolic acidosis can occur with reductions in uteroplacental blood flow, depending on the magnitude and duration.26 However, the relationship between uterine blood flow and oxygen transfer appears nonlinear and suggests that uteroplacental blood flow can decrease by as much as 50% for limited periods before fetal oxygen uptake decreases and metabolic acidosis occurs.24
Studies in sheep have shown that although uterine blood flow varies over a wide range, fetal oxygen uptake remains relatively constant, suggesting that the efficiency of oxygen extraction is greater when perfusion decreases.27 Using an inflatable balloon occluder around the terminal aorta to reduce uterine blood flow in sheep, Wilkening and Meschia24 found that at high levels of oxygen delivery, fetal oxygen uptake was not significantly affected by variations in uterine blood flow; moreover, fetal oxygen uptake became flow dependent only when uterine oxygen delivery was reduced to less than half the baseline value. Boyle et al.,28 investigating the effects of acute uterine arterial embolization with microspheres in sheep, found a linear decrease in fetal aortic oxygen tension as uterine blood flow decreased. However, uterine oxygen consumption did not decrease and fetal hydrogen ion concentration did not increase until uterine blood flow had decreased to approximately 50% of the baseline value. As uterine blood flow diminished, a reduction in uterine venous oxygen content and a greater arteriovenous oxygen content difference were observed, indicating an increase in oxygen extraction. Gu et al.29 reported comparable findings with the compression of the common uterine artery by an inflatable occluder in sheep.
Although the preceding experiments were conducted in sheep, the same principles may apply to humans. The human placenta, like the sheep placenta, is a relatively inefficient oxygen exchanger. Thus, in humans and sheep, the transfer rate of oxygen is affected less by decreases in placental perfusion than the transfer rate in animals with more efficient placentas, such as the rabbit and guinea pig. Of interest, this difference may afford some protection in humans, because alterations in placental perfusion in animals with more efficient placentas frequently result in spontaneous abortion.30 Animal data would also suggest the presence of a significant physiologic buffer that protects the fetus during transient fluctuations in uteroplacental perfusion (e.g., changes in endogenous vasoconstrictor levels, uterine contractions, and parturition).31 This may partially explain why clinical studies have failed to demonstrate fetal acidosis when alpha-adrenergic agonists are used to maintain maternal blood pressure during neuraxial anesthesia,32 despite experimental data showing that these agents reduce uteroplacental perfusion in laboratory animals.33 These observations are based on an assumption of normal physiology; the presence of pathology likely diminishes any margin of safety.
Changes during Parturition
With the onset of the uterine contractions of labor, uteroplacental perfusion undergoes cyclical changes. During uterine contractions, a decrease in perfusion occurs that is inversely related to the strength of the contraction and the increase in intrauterine pressure.31 Conversely, during uterine relaxation, there is a period of hyperemia when perfusion is increased. Placental perfusion is believed to be more sensitive to these contraction-induced changes than myometrial or endometrial blood flow.34 Within the first few hours of parturition, uterine blood flow in sheep decreases on average by 50% or more, although there is notable inter-individual variation.35
Clinical Determinants of Uterine Blood Flow
In the acute setting, uterine blood flow is related to perfusion pressure (the difference between uterine arterial pressure and uterine venous pressure) and vascular resistance, as represented in the following equation:
 (1)
(1)
Thus, there are several ways that uterine blood flow can decrease (Box 3-1). First, uterine blood flow may decline with reductions in perfusion pressure because of decreased uterine arterial pressure—for example, during systemic hypotension from hemorrhage, aortocaval compression, or sympathetic blockade during neuraxial anesthesia. Second, uterine blood flow may decline with reductions in perfusion pressure caused by increased uterine venous pressure—for example, with vena caval compression, increased intrauterine pressure during uterine contractions, drug effects (e.g., oxytocin, cocaine), and Valsalva maneuvers that accompany maternal expulsive efforts during the second stage of labor. Third, uterine blood flow may decline because of increased uterine vascular resistance, which may be caused by a number of factors, including endogenous vasoconstrictors that are released in response to stress, exogenous vasoconstrictors, and compression of endometrial spiral arterioles with uterine contractions.34
Mechanisms of Vascular Changes and Regulation
Vascular Changes during Pregnancy
Because mean arterial pressure decreases slightly during pregnancy, the increase in uteroplacental blood flow is dependent on a substantial decrease in uterine vascular resistance (Figure 3-4), together with increased cardiac output and intravascular volume. The main factors contributing to the decrease in vascular resistance include vascular remodeling, changes in vascular reactivity, and the development of the widely dilated placental circulation.
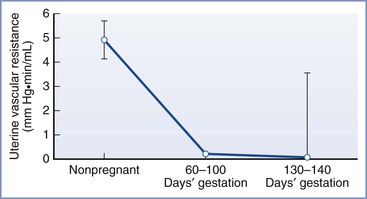
FIGURE 3-4 Changes in uterine vascular resistance with gestation. Data are mean ± SE. (Adapted from Rosenfeld CR. Distribution of cardiac output in ovine pregnancy. Am J Physiol 1977; 232:H231-5.)
Vascular remodeling of arteries in the uterus during pregnancy is believed to include increases in both vessel diameter and vessel length. In humans, both vessel lengthening and straightening of coiled vessels may occur.12 According to Poiseuille’s law, vascular resistance is decreased in proportion to the fourth power of the radius, whereas resistance is increased in proportion to the first power of vessel length; as such, the effects of changes in vessel diameter dominate, resulting in an overall decrease in resistance. Palmer et al.,13 using serial Doppler studies during pregnancy, observed that uterine artery diameter is doubled by 21 weeks’ gestation, whereas there is no change in the diameter of the common iliac or external iliac arteries. These investigators also showed that uterine artery mean flow velocity increased progressively during pregnancy to values eight times greater than those of nonpregnancy. In parallel with arterial changes, there is also structural remodeling of uterine veins in pregnancy. This includes increases in diameter and distensibility and decreases in elastin content.36 Although blood viscosity decreases during pregnancy and also contributes to reduced uterine vascular resistance, this is considered a relatively minor effect compared with vascular changes.37
Changes in vascular reactivity during pregnancy include a vasodilatory response that is meditated at endothelial and vascular smooth muscle levels.38 The growth of the placenta creates a low-resistance vascular pathway by eliminating the intramyometrial microcirculation and creating an intervillous space.39 This has functional characteristics of an arteriovenous shunt.15
The mechanisms underlying the vascular changes during pregnancy are incompletely understood. Contributing factors include steroid hormones, decreased response to vasoconstrictors, endothelium-derived vasodilators, increased shear stress, and venoarterial exchange.
Steroid Hormones
Steroids play an integral role in the development and regulation of the uteroplacental circulation. Estrogen and progesterone are especially important, and there is evidence that cortisol may also contribute.
Estrogen has a fundamental role in the short- and long-term uterine vascular changes during pregnancy. Plasma concentrations of estrogen, initially derived from the ovaries and later predominantly from the placenta, rise concomitantly with the increase in uterine blood flow during pregnancy. Exogenously administered estrogen causes uterine vasodilation and a marked rise in uterine blood flow, independent of systemic effects.40 Angiogenic and vasodilatory effects of estrogen are meditated via estrogen receptors ER-α and ER-β, which are structurally and functionally distinct. The majority of these receptors are located in the nucleus and mediate genomic effects by regulating transcription of genes that are particularly responsible for the longer-term uterine angiogenic responses. There are also membrane receptors that mediate nongenomic effects by up-regulating endothelial production of nitric oxide through the activation of endothelial nitric oxide synthase (eNOS) and the augmentation of eNOS protein expression.41
Progesterone modulates the effect of estrogen on uterine blood flow. In a nonpregnant sheep model, exogenous progesterone administered alone had no uterine vasodilatory effect but had an inhibitory effect when combined with estrogen.38 Progesterone down-regulates expression of estrogen receptors.42 An increase in the estrogen-progesterone ratio parallels the increase in uterine blood flow in late pregnancy in many species.43
Plasma cortisol levels approximately double during pregnancy. Cortisol has both systemic and local effects on uterine blood flow. Systemically, cortisol contributes to regulation of uterine blood flow by increasing plasma volume. Although cortisol is believed to decrease eNOS protein expression and decrease nitric oxide release, it potentiates the response to vasoconstrictor agents including angiotensin II, vasopressin, and norepinephrine. Attenuation of these effects occurs during pregnancy.38
Decreased Response to Vasoconstrictors
In pregnancy, there is a generalized reduction in response to endogenous and exogenous vasoconstrictors, including angiotensin II, endothelin, thromboxane, epinephrine, norepinephrine, phenylephrine, serotonin, thromboxane, and arginine vasopressin.44–46 The relative refractoriness of the systemic and uterine circulations varies for different agents, which has important implications for the regulation and maintenance of uteroplacental blood flow.
During pregnancy, concentrations of angiotensin II in maternal blood are increased twofold to threefold47; however, the vasopressor response to angiotensin II is attenuated.48 This refractoriness is diminished in patients in whom preeclampsia develops.48 The uterine circulation is less responsive to angiotensin II than the systemic circulation. Thus, infusion of physiologic doses of angiotensin II has been shown to have minimal effect on uteroplacental blood flow while increasing systemic blood pressure.49 The difference in sensitivity of the uterine and systemic circulations to angiotensin II is considered an important physiologic adaptation during pregnancy that contributes to the redistribution of cardiac output, the increase in uterine blood flow, and possibly the maintenance of uterine blood flow during normal fluctuations in blood pressure.50
Sensitivity to vasoconstrictors such as epinephrine, norepinephrine, and phenylephrine is attenuated during pregnancy.51 However, in contrast to the responses to angiotensin II, the uterine circulation is more responsive to these agents than the systemic circulation.51 Thus, during hemorrhage or other major stresses that result in large catecholamine release, it is unlikely that uteroplacental perfusion will be preferentially preserved above essential maternal perfusion.52
The mechanism underlying the difference in vascular sensitivity between the uterine and systemic circulations is unclear, but distribution of receptor subtypes is believed to be important.53 There are two distinct subtypes of angiotensin II receptors: AT1R and AT2R. In most tissues, including systemic vascular smooth muscle, AT1R receptors are predominant and mediate vasoconstriction. However, AT2R receptors, which do not mediate smooth muscle contraction, account for 75% to 90% of angiotensin II binding in uterine artery and myometrium.54,55
Vasodilators
The greater synthesis and higher circulating concentrations of endothelial-derived vasodilators during pregnancy are believed to modulate systemic and uterine vascular responses to angiotensin II and other vasoconstrictors.56 Uterine vascular production of prostacyclin is greater than systemic vascular production, which probably contributes to maintaining uteroplacental blood flow in opposition to the effects of circulating vasoconstrictors.57 An enhanced response to angiotensin II during pregnancy has been demonstrated with the systemic and local infusion of indomethacin (which blocks prostacyclin production).58 However, inhibition of prostaglandin synthesis by an infusion of indomethacin induces only a transient decrease in uteroplacental blood flow, indicating that uteroplacental blood flow is not solely dependent on the continued production of prostacyclin.56
Nitric oxide is synthesized from arginine in vascular endothelial cells and stimulates soluble guanylate cyclase in vascular smooth muscle, resulting in vascular relaxation through increases in cyclic guanosine monophosphate. Synthesis of nitric oxide is an important mechanism underlying changes in systemic and uterine vascular resistance, attenuated responses to vasoconstrictors, and vascular effects of estrogen during pregnancy.59 During pregnancy, uterine arteries have increased eNOS activity, higher levels of eNOS messenger ribonucleic acid and eNOS protein, and increased biosynthesis of nitric oxide and cyclic guanosine monophosphate.59,60 Removal of the vascular endothelium diminishes or eliminates the refractoriness of the uterine artery to vasoconstrictors45 and inhibition of nitric oxide synthesis by N-nitro-L-arginine methyl ester (L-NAME) decreases uterine blood flow and also reverses refractoriness to vasoconstrictors.61 Long-term inhibition of nitric oxide synthase causes hypertension and fetal growth restriction in rats.62
Other Vasoactive Substances
Atrial and brain natriuretic peptides attenuate the response to angiotensin II, and intravenous infusion of atrial natriuretic peptide reduces blood pressure while increasing uterine blood flow in preeclamptic women.63 Protein kinase C activity is decreased in uterine, but not systemic, arteries of pregnant sheep and may cause vasodilation and an increase in uterine blood flow; this may have a regulatory effect on local ovarian and placental estrogen production.43 Studies in rats have shown a decrease in endogenous endothelin-dependent vasoconstrictor tone in uteroplacental vessels, which may contribute to the increase in placental blood flow in late gestation.64 Uterine vascular resistance in early pregnancy may be increased by relaxin, which may have a role in modulating the effects of estrogen and progesterone.65
Shear Stress
Shear stress, the frictional forces on the vessel wall from blood flow, is believed to be an important stimulus for uteroplacental vasodilation and remodeling.66 The reduction in downstream resistance resulting from the formation of the placenta would be expected to increase the upstream flow velocity and thus shear stress.39 Nitric oxide is considered an important mediator of this effect because increases in eNOS expression and nitric oxide production are witnessed with shear stress and because stripping the endothelium or pretreatment with L-NAME reduces or abolishes flow-induced vasodilation.66 Studies in vitro have shown that shear stress also increases endothelial production of prostacyclin.
Venoarterial Signaling
It has been postulated that growth factors or signal substances secreted by the placenta and/or myometrium could pass from uterine veins to adjacent uterine arteries; this may provide a system whereby the uterus and placenta regulate their own perfusion.39 Possible candidates for signal substances include vascular endothelial growth factor and placental growth factor. Confirmation of whether this mechanism is important in humans is awaited.
Methods of Measurement of Uteroplacental Blood Flow
Many techniques have been used to measure uteroplacental blood flow in animals and humans. The approaches used in different studies have varied according to the nature of the experimental question, the existing state of technology, and ethical considerations and limitations. All methods have an inherent potential for error.
Many past studies of uterine artery flow have measured flow in only one uterine artery, which may not be an accurate representation of total flow, depending on the location of the placenta (see earlier discussion). The parameter of greatest clinical interest is placental perfusion, but this is not always differentiated from total uterine blood flow, from which it may vary independently. However, in most circumstances, the measurement of intervillous blood flow provides a close approximation of functional placental blood flow. Ovarian arterial blood flow is generally not measured, although studies in primates suggest it may contribute as much as one sixth of placental perfusion.67
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


