Chapter 5
Fetal Physiology
Mieke Soens MD, Lawrence C. Tsen MD
Chapter Outline
Fetal life in utero differs significantly from postnatal life. The fetus relies completely on the mother and the placenta for basic metabolic needs such as nutrient delivery, gas exchange, acid-base balance, and electrolyte homeostasis. During gestation, the fetus gradually assumes the responsibility for many of the vital physiologic functions that must be assumed after the abrupt transition to physiologic independence at birth. Knowledge of fetal physiology, and the timing associated with these developmental changes, is necessary for the optimal provision of analgesia and anesthesia during pregnancy and childbirth.
Fetal Environment
Amniotic Fluid
The fetus is surrounded by amniotic fluid, a complex fluid that changes as the pregnancy progresses. Amniotic fluid serves a number of vital roles, including the facilitation of fetal growth, the provision of a microgravity environment that cushions the fetus, and the generation of a defense mechanism against invading microbes.1,2 The formation and maintenance of amniotic fluid is an intricate process that depends on fetal maturation and maternal hydration, hormonal status, and uteroplacental perfusion.
Amniotic fluid during early embryogenesis is principally derived from maternal plasma by the passage of water and solutes through fetal membranous and placental layers. Between 10 and 20 weeks’ gestation, the volume of amniotic fluid increases in a predictable and linear manner from approximately 25 mL to 400 mL. During this period, the composition of amniotic fluid is similar to fetal extracellular fluid, owing to the absence of keratin in the fetal skin. After this period, the volume of amniotic fluid is a function of production, from fetal urine (600 to 1200 mL/day near term) and respiratory tract secretions (60 to 100 mL/kg fetal body weight/day), and removal through fetal swallowing (200 to 250 mL/kg fetal body weight/day).3 Amniotic fluid volume is also influenced by intramembranous (between amniotic fluid and fetal blood within the placenta) and transmembranous (between amniotic fluid and maternal blood within the uterus) pathways in both physiologic and pathophysiologic states.4 Finally, the status of maternal hydration and the amount of decidual prolactin may alter the transfer of amniotic fluid through fetal and maternal tissues.
The composition of amniotic fluid undergoes more marked variation than its volume.5,6 Keratinization of the fetal skin is complete by 25 weeks’ gestation and decreases the permeability of fetal tissues to water and solutes. The impact of this process, coupled with the ability of the fetal kidneys to produce urine, results in increased amniotic fluid concentrations of urea and creatinine, decreased concentrations of sodium and chloride, and reduced osmolality. A variety of carbohydrates, proteins, lipids, electrolytes, enzymes, and hormones, which vary in concentration depending on the gestational age, are also present; some of these elements, particularly the amino acids taurine, glutamine, and arginine, serve a nutritive function for mitotic cells involved in trophoblastic growth and placental angiogenesis.1 An abundance of growth factors are found in amniotic fluid, including epidermal growth factor, transforming growth factor-alpha, transforming growth factor-beta 1, insulin-like growth factor-1, erythropoietin, and granulocyte colony-stimulating factor; many of these growth factors play an important role in fetal intestinal development.1,7
Antimicrobial defenses within the amniotic fluid are primarily composed of humoral mediators such as alpha-defensins, which are released from neutrophils, especially in the setting of preterm labor and/or chorioamnionitis. Other humoral mediators include lactoferrin, calprotectin, leukocyte protease inhibitor, and cathelicidin, which have significant activity against bacteria, viruses, and fungi.8–10 Cellular mediators of the immune response are poorly characterized in amniotic fluid, and it remains unclear if the macrophages that are present serve a scavenging or an antimicrobial role. Neutrophils are usually absent from the amniotic fluid of a healthy fetus, and their presence typically signifies an inflammatory or infectious process.1
Biochemical and cellular analyses of amniotic fluid provide valuable information on chromosomal abnormalities, neural tube defects, prenatal infections, and most inborn errors of metabolism.11,12 Several amniotic fluid−based indices, including the lecithin-sphingomyelin ratio, the phosphatidylglycerol level, and the lamellar body count, are commonly used to assess fetal lung maturity.13,14 Bilirubin levels can be determined by measuring the optical density of amniotic fluid, which assists in the monitoring of fetal hemolysis. Estimation of the amniotic fluid levels of S100-β (a protein released from injured astrocytes) and cell-free fetal nucleic acids may serve as early screening tests for perinatal neurologic damage and fetal development, respectively.15,16 Finally, amniotic fluid is a valuable reservoir for cell types of multiple lineages at different maturational ages; approximately 1% of these cells are pluripotent, thereby representing a novel source of stem cells.17,18
Oxygen Supply and Transport
The fetus has almost no oxygen reserve and thus depends on maternal sources of oxygen delivery. Oxygen is an essential substrate for cell survival, because it is the final electron acceptor in the electron transport chain. When oxygen is scarce, the electron transport chain is compromised, resulting in decreased oxidative phosphorylation and adenosine triphosphate (ATP) production.19 Hypoxia ensues when the demand for oxygen exceeds the available supply, and it occurs more frequently in the presence of low oxygen tensions. In adult tissues, hypoxia occurs at oxygen tensions less than 20 mm Hg (normal, 40 mm Hg). By contrast, in fetal tissues, hypoxia occurs at oxygen tensions less than 17 mm Hg (normal, 20 to 25 mm Hg).20,21 This implies that fetal development occurs in an environment that exhibits a smaller margin of safety before reaching a state of oxygen insufficiency and highlights the importance of ensuring fetal oxygen delivery through the maintenance of adequate uteroplacental perfusion and fetal cardiac output. Ultimately, oxygenation of fetal tissues depends principally on the partial pressure of oxygen gradient between maternal and fetal blood as well as on the difference in the types of hemoglobin that exist in maternal and fetal blood.
Placental oxygen concentrations change with gestation. In early pregnancy, the placental intervillous space is free of maternal blood cells, thereby requiring the embryo to rely on endometrial secretions and maternal plasma for its energy requirements.22,23 The first trimester placenta has (1) an oxygen partial pressure (PO2) of approximately 20 mm Hg; (2) only a few capillaries, which are located mainly in the center of the mesenchymal core; and (3) a trophoblastic layer that is twice the thickness of that in the second trimester.24 Moreover, the fetal red blood cells are nucleated and the exocoelomic cavity does not contain an oxygen transport system, but rather it contains antioxidant molecules. These anatomic features, which limit the transfer of oxygen and the creation of free radicals, protect the highly sensitive embryo from the effects of oxidative stress.25 At the end of the first trimester, an exponential increase in fetal growth creates significant demands for oxygen and nutrients (Figure 5-1). In response, cytotrophoblastic cells interact with the smooth muscle of maternal spiral arteries, resulting in vessel dilation (Figure 5-2). This allows oxygen-rich maternal blood to flow to the placenta.26
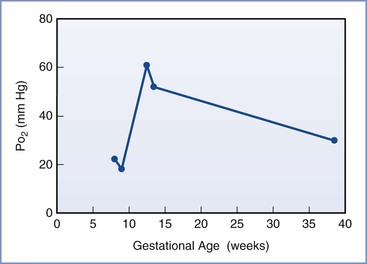
FIGURE 5-1 The mean oxygen partial pressure (PO2) throughout gestation in the human intervillous space. (Data from Jauniaux E, Kiserud T, Ozturk O, et al. Amniotic gas values and acid-base status during acute maternal hyperoxemia and hypoxemia in the early fetal sheep. Am J Obstet Gynecol 2000; 182:661-5; Rodesch F, Simon P, Donner C, Jauniaux E. Oxygen measurements in endometrial and trophoblastic tissues during early pregnancy. Obstet Gynecol 1992; 80:283-5; and Schaaps JP, Tsatsaris V, Goffin F, et al. Shunting the intervillous space: new concepts in human uteroplacental vascularization. Am J Obstet Gynecol 2005; 192:323-32.)
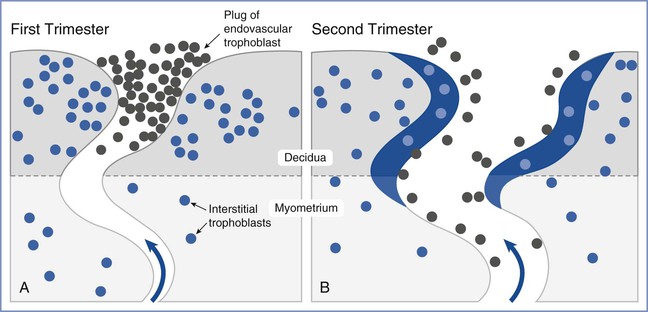
FIGURE 5-2 Invasion and remodeling of the spiral arteries by endovascular and interstitial extravillous trophoblasts. A, In the first trimester, the terminal portion of the spiral artery is blocked by a plug of endovascular trophoblast. Early placental and embryonic development occurs in a state of low oxygen tension, and nutrition at this early stage is derived from secretions from maternal endometrial glands. B, After 10 to 12 weeks’ gestation, the endovascular trophoblast plug dissolves and the endovascular trophoblast migrates into the myometrium, replacing endothelial cells, which undergo apoptosis. Maternal blood is now able to enter the intervillous space, the oxygen tension increases to 60 mm Hg, and nutrition changes from histotrophic to hemotrophic. (Modified from Pijnenborg R, Vercruysse L, Hanssens M. The uterine spiral arteries in human pregnancy: facts and controversies. Placenta 2006; 27:939-58.)
The placenta acts as both a conduit and consumer of oxygen. The placenta is metabolically active and performs important roles in carbohydrate and amino acid metabolism, protein synthesis, and substrate transport. Almost 40% of the oxygen delivered to the pregnant uterus is needed to support the metabolic processes of the placenta.27 During periods of hypoxia, the placenta appears to alter its metabolism to diminish its consumption of oxygen, most likely by increasing glycolysis.28,29 This process can maintain fetal oxygen supply but, if ongoing, may result in fetal growth restriction (also known as intrauterine growth restriction). When the oxygen supply is compromised, the fetus shunts blood flow from peripheral tissues to vital organs (see later discussion), converts to greater use of anaerobic pathways, and undergoes an induction of gene expression that enables improved survival in a low-oxygen environment.19 The presence of fetal hemoglobin (hemoglobin F), with its greater affinity for oxygen than adult hemoglobin (see later discussion), and a hemoglobin concentration higher than that of adults (approximately 18 g/dL) result in a fetal arterial blood oxygen content that is only marginally lower than that in the adult, despite a lower oxygen tension.30
Glucose and Lactate Metabolism
Under normal conditions, gluconeogenesis does not occur to any significant extent in mammalian fetuses; the only source of glucose is that which is transferred across the placenta.31 Fetal glucose concentrations are linearly related to maternal concentrations over a range of 3 to 5 mmol/L (54 to 90 mg/dL; Figure 5-3); studies in isolated placentas suggest that this relationship continues up to a glucose concentration of 20 mmol/L (360 mg/dL).32 The placenta uses the majority of glucose delivered to the uterus for oxidation, glycogen storage, and conversion to lactate, with the remainder being transferred to the umbilical venous blood by facilitated, carrier-mediated diffusion. The amount of glucose supplied to the fetus appears more than adequate during normal conditions; ovine uterine blood flow must be reduced by greater than 50% before a decrease in fetal glucose uptake or fetal arterial glucose concentration is observed.33,34
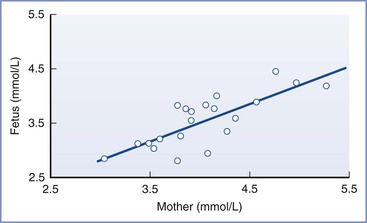
FIGURE 5-3 The linear relationship between maternal and fetal blood glucose concentrations during the third trimester. Fetal blood was obtained by percutaneous umbilical cord blood sampling. (From Kalhan SC. Metabolism of glucose and methods of investigation in the fetus and newborn. In Polin RA, Fox WW, editors. Fetal and Neonatal Physiology. Vol I. Philadelphia, WB Saunders, 1992:477-88.)
The umbilical cord blood glucose uptake is approximately 5 mg/kg/min at normal maternal arterial plasma glucose concentrations.35 Because the umbilical glucose/oxygen quotient varies from approximately 0.5 in sheep36 to 0.8 in human fetuses during labor,37 it is assumed that substrates other than glucose are used to support fetal oxidative metabolism; it is estimated that lactate and amino acids each provides approximately 25% of the total fetal energy requirements.38,39
Lactate is produced even in well-oxygenated fetal lambs, with total lactate production being approximately 4 mg/kg/min.40 Although the exact origin of fetal lactate is unclear, skeletal muscles and bones have been identified as sources of lactate production under resting conditions. Lactate production increases during episodes of acute hypoxemia, although this response may be blunted in fetuses previously exposed to oxidative stress.41 Lactate consumption occurs in the fetal myocardium and liver.42 Short-term exogenous lactate infusion in fetal lambs (sufficient to lower the pH to 7.20) results in transient fetal bradycardia and increased fetal breathing movements but no other adverse effects.43
Amino Acid and Lipid Metabolism
The fetus uses amino acids for protein synthesis, growth, and oxidation. Most maternal-to-fetal amino acid transfer occurs against a concentration gradient and involves energy-dependent transfer mechanisms. Under conditions in which fetal aerobic metabolism is decreased, amino acid uptake by the placenta and fetus may be reduced because it involves an expenditure of energy. Hypoxia results in a large reduction in nitrogen uptake in fetal lambs.44 During maternal fasting, fetal amino acid uptake does not change; however, enhanced fetal proteolysis may occur, which subsequently results in amino acid oxidation or gluconeogenesis.
Lipid products are transferred from the mother to the fetus. The fetus requires free fatty acids for growth, brain development, and the deposition of body fat for postnatal life. Fatty acids are transferred across the placenta by simple diffusion. Ketones are also transferred by simple diffusion; in humans, the maternal/fetal ketone ratio is approximately 2.0.45 The fetus can use ketones as lipogenic substrates or as energy substrates in the brain, kidney, heart, liver, and placenta.46 Beta-hydroxybutyrate (fatty acid) metabolism can occur in the placenta, brain, and liver during episodes of fetal hypoglycemia that result from maternal fasting.46 Cholesterol synthesis or free cholesterol diffusion does not appear to occur in the placenta.47 However, there is a significant correlation between maternal and fetal concentrations of lipoprotein(a), implying that diffusion of lipoprotein(a) may occur.47
Thermoregulation
Intrauterine fetal temperature largely depends on maternal temperature. However, owing to the high metabolic rate in the fetus, the net flow of heat is from the fetus to the mother. When compared with the mother during the third trimester, the fetus produces approximately twice as much heat (on a weight-adjusted basis) and maintains a temperature 0.5° C higher.48,49 This maternal-fetal difference in temperature remains relatively constant and is referred to as the “heat clamp.”50
The placental circulation is responsible for approximately 85% of the heat exchange between the mother and fetus. The remaining 15% is dissipated through the fetal skin and transferred through the amniotic fluid and the uterine wall to the maternal abdomen.51 As a consequence, fetal temperature may be rapidly affected by changes in umbilical blood flow; fetal temperatures rise quickly on occlusion of umbilical blood flow in both baboons and sheep.52,53 In humans, fetal temperatures increase during uterine contractions, which may be a result of intermittent obstruction of umbilical cord blood flow.54 Whether this rise in fetal temperature contributes to acute hypoxic-ischemic brain damage in the setting of umbilical cord prolapse is currently unknown. However, relatively small increases in temperature increase the sensitivity of the fetal brain to hypoxic injury (see Chapter 10).55
Although the fetus generates heat through high metabolic activity, the ability of the fetus to generate heat through thermogenic mechanisms is not developed until the end of gestation and is largely inactive in utero. Newborns are at high risk for rapid heat loss due to amniotic fluid evaporation and a sudden decrease in ambient temperature.49 They are not capable of significant heat production through shivering owing to their small muscle mass. As a consequence, nonshivering thermogenesis plays an important role in maintaining neonatal temperature. Nonshivering thermogenesis occurs in brown adipose tissue, which is unique from other adipocytes owing to the significant presence of mitochondria, fat vacuoles, sympathetic innervation, and blood vessels. In the mitochondria of brown adipose tissue, ATP production is uncoupled from the oxidative process, resulting in an increase in heat production and oxygen consumption.56 Nonshivering thermogenesis is inhibited in utero, most likely owing to the presence of adenosine and prostaglandin E2, which have strong antilipolytic actions on brown tissue.57–59 Inadequate oxygen levels and low levels of intrauterine catecholamines and thyroid hormones may also inhibit nonshivering thermogenesis. The inhibition of nonshivering thermogenesis is believed to be beneficial to the health of the fetus, in that it allows for conservation of fetal oxygenation and accumulation of brown adipose tissue.50
Fetal Cardiovascular System
The cardiovascular system is one of the first functional organ systems in the developing fetus. The morphologic development of the human heart, from its first appearance as a heart tube to its development as a four-chambered structure, occurs between 20 and 44 days’ gestation. Even before the development of the four-chambered heart, the valveless heart tube generates unidirectional flow, typically around 21 days’ gestation.
Circulatory Pattern
Fetal circulation differs significantly from the postnatal circulation. The fetal cardiovascular system is anatomically arranged in such a way as to allow blood to bypass the lungs and provide maximal perfusion of the placenta, where gas and nutrient exchange occur. The fetal systemic circulation receives cardiac output from both the left and the right ventricle, with the ventricles working in parallel. In contrast, during postnatal life, the left and right circulations are separated and the ventricles work in series.
Fetal blood flow is characterized by three anatomic communications between the left and right circulations: the ductus venosus, the foramen ovale, and the ductus arteriosus (Figure 5-4). Oxygenated blood travels from the placenta through the umbilical vein to the ductus venosus, which connects the umbilical vein with the inferior vena cava, thus bypassing the portal circulation and the liver. At mid gestation, approximately 30% of the umbilical venous blood is shunted through the ductus venosus; from 30 to 40 weeks’ gestation, this fraction decreases to approximately 20%, although a significant increase can occur in response to hypoxia (see later discussion).60 Once in the right atrium, oxygenated blood preferentially flows through the foramen ovale to the left atrium and left ventricle, before entering the aorta and the systemic circulation. This mechanism ensures the delivery of well-oxygenated blood to the brain and the heart, which are the two organs with the highest oxygen requirements. The preferential shunting of ductus venosus blood through the foramen ovale into the left atrium is related to the umbilical venous pressure and the portocaval pressure gradient.
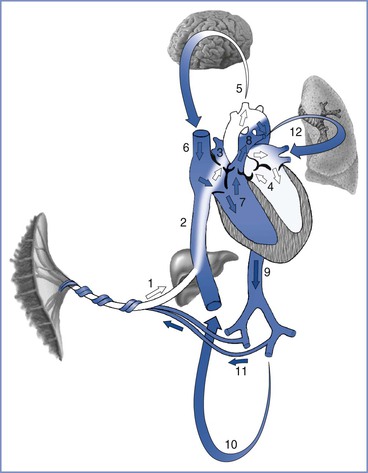
FIGURE 5-4 Oxygenated blood leaves the placenta via the fetal umbilical vein (1), enters the liver where flow divides between the portal sinus and the ductus venosus, and then empties into the inferior vena cava (2). Inside the fetal heart, blood enters the right atrium, where most of the blood is directed through the foramen ovale (3) into the left atrium and ventricle (4), and then enters the aorta. Blood is then sent to the brain (5) and myocardium, ensuring that these cells receive the highest oxygen content available. Deoxygenated blood returning from the lower extremities and the superior vena cava (6) is preferentially directed into the right ventricle (7) and pulmonary trunk. The majority of blood passes through the ductus arteriosus (8) into the descending aorta (9), which in turn supplies the lower extremities (10) and the hypogastric arteries (11). Blood returns to the placenta via the umbilical arteries for gas and nutrient exchange. A small amount of blood from the pulmonary trunk travels through the pulmonary arteries (12) to perfuse the lungs. Arrows in this figure depict the direction and oxygen content [white (oxygenated), blue (deoxygenated)] of the blood in circulation. (Drawing by Naveen Nathan, MD, Northwestern University Feinberg School of Medicine, Chicago, IL.)
Deoxygenated blood from the head and upper extremities enters the right atrium through the superior vena cava and is preferentially directed into the right ventricle and the pulmonary artery. Because fetal pulmonary vascular resistance is higher than systemic vascular resistance, the majority of pulmonary artery blood flow crosses the ductus arteriosus into the descending aorta, which in turn supplies the lower extremities and hypogastric arteries. Deoxygenated blood returns to the placenta via the umbilical arteries for gas and nutrient exchange; only a small percentage travels through the lungs into the left atrium, the left ventricle, and the ascending aorta.
At birth, the fetus undergoes a significant and abrupt transition to a state of physiologic independence (see Chapter 9). Clamping of the umbilical cord results in a sudden increase in systemic vascular resistance, whereas expansion of the lungs and an increased alveolar oxygen tension result in decreased pulmonary vascular resistance. This allows for greater blood flow through the lungs, resulting in a decrease in right atrial pressure and an increase in left atrial pressure, ultimately leading to the functional closure of both the foramen ovale and the ductus arteriosus.
Blood Volume
Human fetal intravascular volume is approximately 110 mL/kg, which is higher than that in postnatal life. However, approximately 25% of this blood volume is contained within the placenta; the blood volume within the fetal body is estimated to be approximately 80 mL/kg.61,62 Fetal intravascular volume is regulated through a complex interplay between the fetal heart, kidneys, and circulation and the placenta.63 The fetus can adapt more quickly to changes in intravascular volume than the adult, owing to higher diffusion rates between fetal compartments.64
Transplacental transfer of water from mother to fetus depends on hydrostatic and osmotic pressures. The hydrostatic pressure is determined by the difference in pressures between the maternal intervillous space or capillaries and the fetal capillaries. The osmotic pressure is mainly determined by the presence of plasma proteins (i.e., colloid osmotic pressure). Transplacental water transfer is further regulated by angiotensin II. Adamson et al.65,66 found that angiotensin II lowered the pressures in fetal placental exchange vessels, thereby promoting fluid transfer from the maternal to the fetal circulation. The production of angiotensin II is under control of the renin-angiotensin-aldosterone system in the fetal kidneys. A reduction in fetal arterial pressure results in an increase in fetal plasma renin activity, which results in subsequent increases in angiotensin I and II. The resulting expansion of intravascular volume augments fetal cardiac output and arterial pressure.
Cardiac Development
During gestation the fetal heart grows quickly and adapts to the continuously changing demands. The fetal myocardium grows primarily through cell division, whereas after delivery, cardiac mass increases as a result of cell enlargement.67 This growth correlates with a pre-birth transition from mononucleated cardiomyocytes, which contribute to heart growth by hyperplasia, to binucleated cardiomyocytes, which contribute to heart growth by hypertrophy.
The number of cardiac myofibrils and the transition in the type of cardiac troponins that are present during prenatal development can alter fetal heart contractility.68 The change from fetal to adult troponin is associated with decreased sensitivity of the contractile apparatus to calcium. A heightened calcium sensitivity is important in the early development of the fetal heart, when the sarcoplasmic reticulum is immature.69 With advancing gestational age, ejection fraction declines but cardiac output (per unit of fetal weight) does not change owing to increasing ventricular volume. The fetal heart rate decreases over the course of gestation from 140 to 150 beats per minute at 18 weeks’ gestation to 120 to 140 beats per minute at term.70,71
Ventricular Responses to Changes in Preload and Afterload
It is unclear whether fetal and adult hearts possess similar responses to preload and afterload. The adult heart responds in accordance to the Frank-Starling curve, which indicates that ventricular distention lengthens the diastolic fibers and results in augmented contractility. A number of studies have indicated that the fetal heart has a limited capacity to increase its stroke volume in response to an increase in preload (e.g., intravenous fluid infusion).72,73 By contrast, other studies have observed that the fetal heart can accommodate increases in preload and afterload in a manner consistent with the Frank-Starling curve.74,75 These seemingly contradictory findings may be partially explained if the fetal heart functions in vivo near the peak of the Frank-Starling curve. However, the left ventricular stroke volume is known to double at birth, which would not be in agreement with this hypothesis. A more plausible explanation is that ventricular constraint, arising from tissues that surround the heart (chest wall, pericardium, and lungs), limits fetal ventricular preload and overall cardiac function in utero. Relief of this constraint at birth, as a result of lung aeration and clearance of liquid from the lungs, may then allow for an increase in left ventricular preload and subsequent stroke volume in the newborn.76
Studies investigating the effects of afterload on fetal ventricular function have observed a significant decrease in right ventricular stroke volume in response to increases in arterial pressure.72 The same phenomenon occurs in the left ventricle, although to a lesser degree. In a study of fetal lambs, in which gradual constriction of the descending aorta was applied, stroke volume was maintained until high mean arterial pressures were achieved, after which decreases were observed. This decrease in stroke volume in the presence of high mean arterial pressure may represent the exhaustion of “preload reserve,” which will typically allow the maintenance of stroke volume in the setting of increased afterload.77
Cardiac Output and Distribution
In postnatal life, the right and left ventricles operate in series and their output is approximately equal; as a consequence, cardiac output is defined through measurements of output from either ventricle. However, in the fetus, the systemic circulation receives blood from both the left and right ventricle in parallel (i.e., the sum of the right and left ventricular outputs, with the exception of a proportion of the right ventricular output that is delivered to the fetal lungs). At mid gestation, the combined ventricular output (CVO) is approximately 210 mL, and it increases to approximately 1900 mL at 38 weeks’ gestation (500 mL/min/kg).73,78,79 During fetal life, the right ventricular volume is greater than the left ventricular volume during both systole and diastole, but stroke volume does not differ significantly between the two ventricles.70
Fetal cardiac output is sensitive to changes in fetal heart rate. As heart rate increases, cardiac output increases. As fetal heart rate decreases, fetal stroke volume increases only slightly, in part because of low fetal myocardial compliance. Although fetal bradycardia results in an extended diastolic filling time, the stiff fetal cardiac ventricles have limited ability to distend. Therefore, fetal bradycardia is associated with a marked drop in fetal cardiac output.
The distribution of the CVO in near-term fetal lambs and resting adult humans is shown in Figure 5-5. The fetal lamb CVO is distributed to the placenta (41%), the bone and skeletal muscle (38%), the gastrointestinal system (6%), the heart (4%), the brain (3%), and the kidneys (2%). In both fetal and adult animals, approximately equal volumes of blood are delivered to oxygen-uptake organs (i.e., the placenta before delivery, the lungs after delivery) and the oxygen-consuming organs.
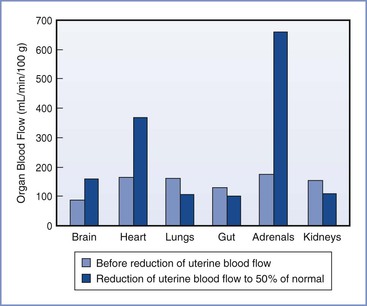
FIGURE 5-5 Redistribution of combined ventricular output in fetal lambs during hypoxemia caused by reduced uterine blood flow. (Modified from Jensen A, Roman C, Rudolph AM. Effects of reducing uterine blood flow on fetal blood flow distribution and oxygen delivery. J Dev Physiol 1991; 15: 309-23.)
The distribution of the CVO changes over the course of gestation and in certain conditions, such as hypoxia and hypovolemia. Interpretation of CVO data should be evaluated with the understanding that significant interspecies differences exist. For example, in humans the fetal lungs receive approximately 20% of CVO, whereas in the fetal lamb the lungs receive 10% or less of CVO. In human fetuses at 10 to 20 weeks’ gestation, the brain receives approximately 15% of CVO,80 but this fraction may be increased during circumstances of decreased placental perfusion, acidosis, and increased PCO2. In the rhesus monkey, the fraction of CVO devoted to cerebral blood flow was observed to increase from 16% to 31% during a hypoxic challenge.81
Fetal Blood Pressure
Fetal blood pressure increases with gestational age. Intracardiac (intraventricular) pressure recordings in the human fetus suggest that systolic pressure increases from 15 to 20 mm Hg at 16 weeks’ gestation to 30 to 40 mm Hg at 28 weeks’ gestation.67 Substantial variation in blood pressure may be observed. The diastolic ventricular pressures undergo similar, albeit slower and smaller increases, from 5 mm Hg or less at 16 to 18 weeks’ gestation to 5 to 15 mm Hg at 19 to 26 weeks’ gestation.67
Regulation of Fetal Circulation
Fetal cardiovascular function continuously adapts to varying metabolic and environmental conditions through regulation by the neurologic and endocrine systems. The predominant form of neuroregulation occurs in response to baroreceptor and chemoreceptor afferent input to the autonomic nervous system and through modulation of myocardial adrenergic receptor activity. Thus, the autonomic nervous system functions to reversibly redirect blood flow and oxygen delivery as required.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


