Race
Pigmentary differences
Black (in the USA: African American or Afro Caribbean)
Large (Stage IV) melanosomes
Eumelanin constitutes majority of pigment production
Closely packed doublet or singlet melanosomes, rare aggregatesa
Larger melanophages (may account in part for greater incidence of melasma and erythema dyschromicum perstans)
UV filtration is in the malpighian layer
Caucasian
Small, aggregated melanosomes
Pheomelanin
Few small melanophages
UV filtration in the stratum corneum
Genetics of Pigmentation
Pigmentation is polygenic with contribution from many types of genes ranging from melanin production, distribution, and dispersion genes to melanoblast migration genes.
Alteration in pigmentation genes produces pigmentary alterations ranging from mild skin tone alterations to complete absence of melanin production.
Pigmentation is polygenic with many different types of genes contributing to the formation of skin tone. Over the last century, many of the studies that discovered genes controlling skin color were investigating pigmentation disorders in humans and animal models. For example, in mice there are greater than 100 genes known to contribute to over 800 phenotypic alleles [21]. With the sequencing of the human genome just over a decade ago, there has been an explosion of new knowledge in this area from studies including comparative genomic and specific allele association studies. Single nucleotide polymorphisms (SNPs) have also been identified in genome-wide association studies that have allowed illumination of genetic variants associated with human pigmentation of the skin, eye, and hair [22]. The most commonly studied genes, TRY, TRP1, P, MATP, MC1R, ASIP, SLC24A5, and MATP, will be described briefly.
The TYR gene encodes for tyrosinase, a copper-dependent enzyme responsible for catalyzing melanin. This gene is mutated in oculocutaneous albinism type 1, with complete or partial loss of gene function (types Ia and Ib, respectively). At present, there are over 100 mutations associated with albinism or skin color dilution [23]. Also contributing to the tyrosinase enzyme complex is TRP1, mutations in which result in oculocutaneous albinism type 3. In individuals of sub-Saharan African heritage with oculocutaneous albinism type 2, mutations in the P gene cause a defective melanocytic transporter protein resulting in light blond or yellow hair, vision problems, and white skin. MATP is a membrane-associated transporter protein associated with OCA type 4 that shows strong selection in European populations.
Another widely studied pigmentation gene is the melanocortin 1 receptor (MC1R, also called alpha melanocyte stimulating hormone) gene, which codes for a G protein-coupled receptor important in melanocytic switching between production of eumelanin and pheomelanin. Loss-of-function mutations of MC1R have been associated with people with red hair and fair skin (autologous to an autosomal recessive trait), but are also seen in up to 30 % of the population and may play a role in lighter skin color [24]. European populations show a higher sequencing diversity of MC1R, which reflects neutral expectations of selection under relaxation of functional constraints especially when compared to sub-Saharan African and other dark-skinned populations, which are thought to be under stronger functional constraints and show a lack of sequencing diversity [25, 26].
ASIP, the agouti signaling protein, acts antagonistically at the MC1R receptor to inhibit the production of both eumelanin and pheomelanin; and the 8818G allele is strongly associated with dark hair, brown eyes, and dark skin [27, 28]. Lastly, a gene contributing to the “lightening” of skin is the “golden” gene (SLC24A5) coding for a melanosomal cation exchanger and responsible for up to 25–38 % of the difference between the European versus African melanin index of the skin [29]. Other genes implicated in pigmentation are seen in the Table 2.2 (Fig. 2.1).
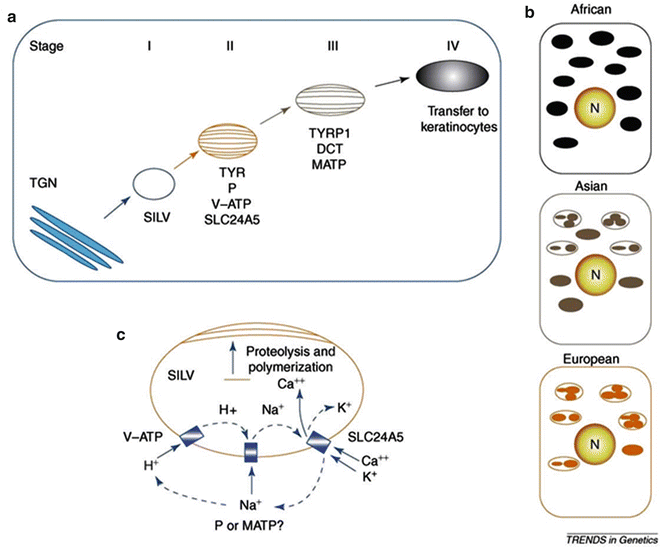
Table 2.2
Genes that contribute to pigmentation
Type | Gene | Associated conditions |
|---|---|---|
Tyrosinase enzyme complex | TYR | Oculocutaneous albinism type I/amelanotic melanoma/vitiligo |
TRP1 | Oculocutaneous albinism type 3/vitiligo | |
Melanosomal proteins | MATP | Oculocutaneous albinism type 4 |
P gene | Oculocutaneous albinism type 2 | |
PMEL/SILV | Juvenile xanthogranuloma/melanoma | |
SLC24A5 | OCA6/Loeys Dietz/patent ductus arteriosus | |
Regulators of melanin synthesis | MC1R/Alpha-MSH | Melanogenesis/eumelanin production/Skin Cancers |
ASIP | Hair pigmentation; skin cancers | |
ATRN | Radioulnar synostosis | |
Transcription factors of melanin production | PAX3 | Waardenburg syndrome, alveolar rhabdomyosarcoma |
MITF | Waardenburg syndrome, types 2 and 2a | |
SOX10 | Waardenburg syndrome, type 4/Nodular melanoma | |
Melanosomal transport proteins | MYO5A | Griscelli syndrome, types I and III; Elejalde syndrome |
MYO7A | Usher syndrome Ib | |
RAB27A | Griscelli syndrome, type II | |
Melanosomal construction/protein routing | CHS1 | Chediak–Higashi syndrome |
HPS1-6 | Hermansky–Pudlak syndrome | |
Developmental ligands controlling melanoblast migration and differentiation | EDN3 | Hirschsprung syndrome/Waardenburg’s syndrome |
KITLG | Familial progressive hyperpigmentation | |
Developmental receptors controlling melanoblast migration and differentiation | KIT | Piebaldism and urticaria pigmentosum/mastocytosis |
Fig. 2.1
Melanosome formation and the role of ion transport in their maturation. (a) The four-stage model of melanosome formation is shown together with key proteins that are necessary for each step of maturation before melanosomes are passed to keratinocytes. (b) Keratinocyte distribution of melanosomes in ethnic populations, note that the melanosomes often form a cap surrounding the nucleus that might have a role in photoprotection. (c) A model for ion transport that is essential to melanosome function. The coupling of H+, Na+ exchange by the V–ATP complex, with possible involvement of the P or MATP proteins, enables SLC24A5 (also known as NCKX5) to transport K+, Ca2+ ions into the melanosome. Ca2+ might have an essential role in activating the proteolytic cleavage of SILV, which polymerizes to form the melanosomal matrix copied with permission from Sturm R. 2006 [30]
Hair
Hair type can be categorized by shape of hair in cross section, curvature or lack thereof of the follicle, density of the hairs, and content of sulfur in the hairs.
Hair type may have racial or ethnic association for the general public.
Hair in Black patients can be more susceptible to illness due to reduced density, less elastic anchorage, and cultural styling practices.
In the literature, research often focuses on grouping hair textures into African, Caucasian, or Asian. These distinctions, though heavily studied and helpful, do not take into account the multitude of the world’s population that may not fall into one of these categories due to inter- and inner-group variation of hair types. A study of 1,442 people from 18 countries revealed 8 different hair types [31]. Hardy classified hair types without incorporating race, but the classifications have not been widely used [32]. Khumalo suggests that race has been used as a proxy for describing hair forms, despite obvious inter-racial variation [33, 34].
Khumalo expressed the need for an easy to use classification of hair forms that is inclusive of multiple hair types. Despite this desire, the most commonly used terminology to describe hair types still utilizes racial classifications.
When cross sections of hair are viewed, Asian hair is round. Caucasian hair is thinner and more elliptical than Asian hair. African hair is often textured, is coiled, and is the most elliptical [35]. On cross section, Black hairs will be flattened. Textured hair and dryness of the scalp and hairs are common in Black hair, due to reduced sebum production/distribution along the hair shaft and reduced water content in the hairs. The hair follicle is expected to be helical or curved, with limited elastic fiber anchorage. Lower hair density is noted in Black patients (0.6 follicular units per square mm vs. 1 follicular unit per square meter in Asians and Caucasians) [36–38].
It is well known that African textured hair may be straightened permanently with chemical relaxers and temporarily with heat, with side effects ranging from frizziness to follicular destruction [39]. Close observation has revealed that the African texture is also noted to change during certain types of illnesses and states of health such as AIDS, rheumatoid arthritis, systemic lupus erythematosus, pulmonary tuberculosis with cachexia, and Behçet’s disease, especially those with anemia of chronic illness, high erythrocyte sedimentation rate, and mild hypocalcemia [40].
In infants, the hair whorl may be hard to note in Black children due to the curl of the hair compounded by the popularity of shaven hairstyles. Microscopy of hair in Black children demonstrates discrete hair packets and curled hairs.
Follicular prominence can be noted in Black adolescents resulting in a light halo near each sebaceous hair follicle of the face. Follicular inflammation is more common in Black children with consequently greater amounts of follicular eczema and folliculocentric allergic contact dermatitis.
Dermal
Fibroblasts are larger in the dermis of Black children contributing to the increased incidence of keloidal lesions in this racial group.
Elastic tissue anchorage of the hair follicle is reduced resulting in greater damage with traction-based hairstyles.
Skin thickness is the same in Blacks and Whites [43], despite the compact nature of the stratum corneum in Blacks [44]. Fibroblasts in Black skin are larger than those in White skin [45]. Elastic tissue anchorage of the hair follicle is reduced in Black patients resulting in greater risk of traction-induced damage.
Keloids result from unbalanced extracellular matrix production and degradation [46]. Hyperactive fibroblasts contribute to keloid formation and are influenced by transforming growth factor beta, epidermal growth factor, mast cells, and decreased collagenase activity [47–49].
Keloid development is influenced by many factors including genetic susceptibility including racial prevalence amongst Blacks, Asians, and Hispanics, family linkage, and HLA associations and corroborated by twin studies. Environmental contributory factors include hormones, wound tension, infection, and foreign body granulomas. Another factor that authors note in practice is the comorbidity of nickel contact allergy, often induced by piercing, as a trigger of keloids secondary to piercing.
Other Considerations
Genetic differences in metabolism and skin structure can affect response to medications.
Awareness of G6PD deficiency, an X-linked recessive enzymatic defect, is needed for practitioners who prescribe dapsone and antimalarial medications, due to the risk of severe hemolysis.
The role of environment on development of skin diseases is especially contributory in the development of atopic dermatitis in developed countries.
Some dermatologic diseases affecting patients with skin of color have been linked to genetic propensity. For example, sarcoid, is associated with specific HLA types in Black patients [50, 51].
Vitiligo is more prominent in children of color, but despite this, no specific linkage genes to race have been identified in Black children. Vitiligo genetics is actually polygenic and multifactorial [52]. On the other hand, OCA2, an autosomal recessive albinism, has a specific gene defect and is the most prevalent autosomal recessive disease among South African Blacks, P protein is defective in OCA2 leading to abnormal tyrosinase enzyme function and defective melanin production [52].
Keloids have long been observed to occur more frequently in skin of color populations, especially in those of African descent. Studies now suggest that certain environmental triggers may spur keloid formation in those who are genetically susceptible [53].
Other pertinent genetically common illnesses in patients of Black or African descent include G6PD deficiency, an X-linked recessive enzymatic defect that affects metabolism of medications such as dapsone and hydroxychloroquine and can result in severe hemolysis with drug administration of these agents. Male patients should be suspected most, but all black patients should be screened prior to usage of these agents as female patients may be homozygous or have low expression based on lyonization.
Sickle cell anemia can confer susceptibility to bacterial infection (e.g., Streptococcus) [54] and is associated with severe hemolysis requiring hospitalization for transfusion in children with G6PD deficiency. Sickle cell carriers may be less prone to malaria, generating the hypothesis as to why sickle cell carriage and disease are more common in patients of African descent [55].
Type II diabetes mellitus is associated with acanthosis nigricans, skin tags, candidal infections, and poor wound healing. In the USA, Black, Native American/Inuit, and Mexican American children are at increased risk. Signs of insulin resistance, especially acanthosis nigricans, are noted in pre-teen years with disease becoming full blown in some cases by the mid-teen years [56, 57].
Black children also have specific reduction in the formation of infantile hemangiomas [58] and lifetime risk of skin cancers [59] (lifetime risk is lower). Collagen vascular diseases are more common from birth, i.e., neonatal lupus through childhood/adolescence when Black children may develop the first features of lupus erythematosus, with specifically increased risk of nephritis [60]. More than 60 % of patients under the age of 20 years with systemic lupus erythematosus will be Black [61].
The effect of the environment/country/place of birth on the disease incidence cannot be ignored, e.g., atopic dermatitis being more common in Afro-caribbeans in London, but relatively less common in Africans on the continent, etc. In addition, differences in the pattern of skin disease exist between races. Henderson et al. found that more than 60 % of all pediatric patients seen at their dermatology clinic had diagnoses of acne (28.6 %), dermatitis (19.4 %), and warts (16.2 %) [62]. But when the patients were further stratified, they found that African-American pediatric patients in their study were most commonly seen for dermatitis (29.0 %), acne (27.5 %), and dermatophytosis (10.2 %), whereas Caucasian children were most commonly seen for acne (29.9 %), warts (22.6 %), and dermatitis (13.1 %).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


