Chapter 557 Thyroid Development and Physiology Stephen LaFranchi Fetal Development The fetal thyroid bilobed shape is recognized by 7 wk of gestation, and characteristic thyroid follicle cell and colloid formation is seen by 10 wk. Thyroglobulin synthesis occurs from 4 wk, iodine trapping occurs by 8-10 wk, and thyroxine (T4) and, to a lesser extent, triiodothyronine (T3) synthesis and secretion occur from 12 wk of gestation. There is evidence that several transcription factors—TTF-1/NKX-2.1, TTF-2 (also termed FOXE1), NKX2.5, and PAX8—are important in thyroid gland morphogenesis and differentiation and possibly also in its caudal migration to its final location. These factors also bind to the promoters of thyroglobulin and thyroid peroxidase genes and so influence thyroid hormone production. Hypothalamic neurons synthesize thyrotropin-releasing hormone (TRH) by 6-8 wk, the pituitary portal vessel system begins development by 8-10 wk, and thyroid-stimulating hormone (TSH) secretion is evident by 12 wk of gestation. Maturation of the hypothalamic-pituitary-thyroid axis occurs over the 2nd half of gestation, but normal feedback relationships are not mature until approximately 3 mo of postnatal life. Another transcription factor, Pit-1, is important for differentiation and growth of thyrotrophs, along with somatotrophs and lactotrophs. Thyroid Physiology The main function of the thyroid gland is to synthesize T4 and T3. The only known physiologic role of iodine (or iodide [I−] in its ionized form) is in the synthesis of these hormones; the recommended dietary allowance of iodine is 30 µg/kg/24 hr for infants, 90-120 µg/24 hr for children, and 150 µg/24 hr for adolescents and adults. The median iodine intake in the United States has decreased by approximately 50% between the 1970s (320 µg/L) and the 1990s (145 µg/L), although at present intake appears to have stabilized. Whatever the chemical form ingested, iodine eventually reaches the thyroid gland as iodide. Thyroid tissue has an avidity for iodide and is able to trap (with a gradient of 100 : 1), transport, and concentrate it in the follicular lumen for synthesis of thyroid hormone. Entry of iodide from the circulation into the thyroid is carried out by the sodium-iodide symporter. Before trapped iodide can react with tyrosine, it must be oxidized; this reaction is catalyzed by thyroidal peroxidase. Dual oxidase maturation factor 2 (DUOXA2) is required to express DUOX2 enzymatic activity, which is required for H2O2 generation, a crucial step in iodide oxidation. The thyroid cells also elaborate a specific thyroprotein, a globulin with approximately 120 tyrosine units (thyroglobulin). Iodination of tyrosine forms monoiodotyrosine and diiodotyrosine; 2 molecules of diiodotyrosine then couple to form 1 molecule of T4, or 1 molecule of diiodotyrosine and 1 of monoiodotyrosine to form T3. Once formed, hormones are stored as thyroglobulin in the lumen of the follicle (colloid) until ready to be delivered to the body cells. Thyroglobulin is a large globular glycoprotein with a molecular weight of about 660,000. T4 and T3 are liberated from thyroglobulin by activation of proteases and peptidases. The metabolic potency of T3 is 3 to 4 times that of T4. In adults, the thyroid produces approximately 100 µg of T4 and 20 µg of T3 daily. Only 20% of circulating T3 is secreted by the thyroid; the remainder is produced by deiodination of T4 in the liver, kidney, and other peripheral tissues by type I 5′-deiodinase. Selenocysteine is the active center of the iodothyronine deiodinases. Thus, selenium indirectly plays a role in normal growth and development. In the pituitary and brain, approximately 80% of required T3 is produced locally from T4 by a different enzyme, type II 5′-deiodinase. The level of T3 in blood is that of T4, but T3 is the physiologically active thyroid hormone. Thyroid hormones increase oxygen consumption, stimulate protein synthesis, influence growth and differentiation, and affect carbohydrate, lipid, and vitamin metabolism. The free hormones enter cells, where T4 may be converted to T3 by deiodination. Monocarboxylate transporter 8 is an active, specific thyroid hormone transporter that facilitates T4 entry into cells. Mutations of the MCT8 are associated with high T3 levels, thyroid hormone resistance, and severe X-linked psychomotor retardation. Intracellular T3 then enters the nucleus, where it binds to thyroid hormone receptors. Thyroid hormone receptors are members of the steroid hormone receptor superfamily that includes glucocorticoids, estrogen, progesterone, vitamin D, and retinoids. Four different isoforms of the thyroid hormone receptor (α1, α2 β1, and β2) are expressed in different tissues; the protein product of the formerly designated c-erb A proto-oncogene (THRA2) is the α2 thyroid hormone receptor in the brain and hypothalamus. Thyroid hormone receptors consist of a ligand-binding domain (binds T3), hinge region, and DNA-binding domain (zinc finger). Binding of T3 activates the thyroid hormone receptor response element, resulting in production of an encoded mRNA and protein synthesis and of secretion specific for the target cell. In this manner, a single hormone, T4, acting through tissue-specific thyroid hormone receptor isoforms and gene-specific thyroid response elements, can produce multiple effects in various tissues. About 70% of the circulating T4 is firmly bound to thyroxine-binding globulin (TBG). Less-important carriers are thyroxine-binding prealbumin, called transthyretin, and albumin. Only 0.03% of T4 Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Related Related posts: Rumination, Pica, and Elimination (Enuresis, Encopresis) Disorders Adolescent Pregnancy Neisseria gonorrhoeae (Gonococcus) Blastomycosis (Blastomyces dermatitidis) Stay updated, free articles. Join our Telegram channel Join Tags: Nelson Textbook of Pediatrics Expert Consult Jun 18, 2016 | Posted by admin in PEDIATRICS | Comments Off on Thyroid Development and Physiology Full access? Get Clinical Tree
Chapter 557 Thyroid Development and Physiology Stephen LaFranchi Fetal Development The fetal thyroid bilobed shape is recognized by 7 wk of gestation, and characteristic thyroid follicle cell and colloid formation is seen by 10 wk. Thyroglobulin synthesis occurs from 4 wk, iodine trapping occurs by 8-10 wk, and thyroxine (T4) and, to a lesser extent, triiodothyronine (T3) synthesis and secretion occur from 12 wk of gestation. There is evidence that several transcription factors—TTF-1/NKX-2.1, TTF-2 (also termed FOXE1), NKX2.5, and PAX8—are important in thyroid gland morphogenesis and differentiation and possibly also in its caudal migration to its final location. These factors also bind to the promoters of thyroglobulin and thyroid peroxidase genes and so influence thyroid hormone production. Hypothalamic neurons synthesize thyrotropin-releasing hormone (TRH) by 6-8 wk, the pituitary portal vessel system begins development by 8-10 wk, and thyroid-stimulating hormone (TSH) secretion is evident by 12 wk of gestation. Maturation of the hypothalamic-pituitary-thyroid axis occurs over the 2nd half of gestation, but normal feedback relationships are not mature until approximately 3 mo of postnatal life. Another transcription factor, Pit-1, is important for differentiation and growth of thyrotrophs, along with somatotrophs and lactotrophs. Thyroid Physiology The main function of the thyroid gland is to synthesize T4 and T3. The only known physiologic role of iodine (or iodide [I−] in its ionized form) is in the synthesis of these hormones; the recommended dietary allowance of iodine is 30 µg/kg/24 hr for infants, 90-120 µg/24 hr for children, and 150 µg/24 hr for adolescents and adults. The median iodine intake in the United States has decreased by approximately 50% between the 1970s (320 µg/L) and the 1990s (145 µg/L), although at present intake appears to have stabilized. Whatever the chemical form ingested, iodine eventually reaches the thyroid gland as iodide. Thyroid tissue has an avidity for iodide and is able to trap (with a gradient of 100 : 1), transport, and concentrate it in the follicular lumen for synthesis of thyroid hormone. Entry of iodide from the circulation into the thyroid is carried out by the sodium-iodide symporter. Before trapped iodide can react with tyrosine, it must be oxidized; this reaction is catalyzed by thyroidal peroxidase. Dual oxidase maturation factor 2 (DUOXA2) is required to express DUOX2 enzymatic activity, which is required for H2O2 generation, a crucial step in iodide oxidation. The thyroid cells also elaborate a specific thyroprotein, a globulin with approximately 120 tyrosine units (thyroglobulin). Iodination of tyrosine forms monoiodotyrosine and diiodotyrosine; 2 molecules of diiodotyrosine then couple to form 1 molecule of T4, or 1 molecule of diiodotyrosine and 1 of monoiodotyrosine to form T3. Once formed, hormones are stored as thyroglobulin in the lumen of the follicle (colloid) until ready to be delivered to the body cells. Thyroglobulin is a large globular glycoprotein with a molecular weight of about 660,000. T4 and T3 are liberated from thyroglobulin by activation of proteases and peptidases. The metabolic potency of T3 is 3 to 4 times that of T4. In adults, the thyroid produces approximately 100 µg of T4 and 20 µg of T3 daily. Only 20% of circulating T3 is secreted by the thyroid; the remainder is produced by deiodination of T4 in the liver, kidney, and other peripheral tissues by type I 5′-deiodinase. Selenocysteine is the active center of the iodothyronine deiodinases. Thus, selenium indirectly plays a role in normal growth and development. In the pituitary and brain, approximately 80% of required T3 is produced locally from T4 by a different enzyme, type II 5′-deiodinase. The level of T3 in blood is that of T4, but T3 is the physiologically active thyroid hormone. Thyroid hormones increase oxygen consumption, stimulate protein synthesis, influence growth and differentiation, and affect carbohydrate, lipid, and vitamin metabolism. The free hormones enter cells, where T4 may be converted to T3 by deiodination. Monocarboxylate transporter 8 is an active, specific thyroid hormone transporter that facilitates T4 entry into cells. Mutations of the MCT8 are associated with high T3 levels, thyroid hormone resistance, and severe X-linked psychomotor retardation. Intracellular T3 then enters the nucleus, where it binds to thyroid hormone receptors. Thyroid hormone receptors are members of the steroid hormone receptor superfamily that includes glucocorticoids, estrogen, progesterone, vitamin D, and retinoids. Four different isoforms of the thyroid hormone receptor (α1, α2 β1, and β2) are expressed in different tissues; the protein product of the formerly designated c-erb A proto-oncogene (THRA2) is the α2 thyroid hormone receptor in the brain and hypothalamus. Thyroid hormone receptors consist of a ligand-binding domain (binds T3), hinge region, and DNA-binding domain (zinc finger). Binding of T3 activates the thyroid hormone receptor response element, resulting in production of an encoded mRNA and protein synthesis and of secretion specific for the target cell. In this manner, a single hormone, T4, acting through tissue-specific thyroid hormone receptor isoforms and gene-specific thyroid response elements, can produce multiple effects in various tissues. About 70% of the circulating T4 is firmly bound to thyroxine-binding globulin (TBG). Less-important carriers are thyroxine-binding prealbumin, called transthyretin, and albumin. Only 0.03% of T4 Only gold members can continue reading. Log In or Register to continue Share this: Share on X (Opens in new window) X Share on Facebook (Opens in new window) Facebook Related Related posts: Rumination, Pica, and Elimination (Enuresis, Encopresis) Disorders Adolescent Pregnancy Neisseria gonorrhoeae (Gonococcus) Blastomycosis (Blastomyces dermatitidis) Stay updated, free articles. Join our Telegram channel Join Tags: Nelson Textbook of Pediatrics Expert Consult Jun 18, 2016 | Posted by admin in PEDIATRICS | Comments Off on Thyroid Development and Physiology Full access? Get Clinical Tree
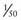 that of T4, but T3 is the physiologically active thyroid hormone.
that of T4, but T3 is the physiologically active thyroid hormone.


