The main stages of follicle growth

Cross-section through an antral, or Graafian, follicle
Selection of a single follicle and oocyte
Folliculogenesis is defined as growth of the follicles from the earliest resting phases through to ovulation. Only the latter stages of this process are dependent on the hormonal changes of the menstrual cycle. The granulosa cells develop receptors for FSH and the theca cells develop receptors for luteinising hormone (LH). Once follicles develop an antrum, they are dependent on FSH for further growth.
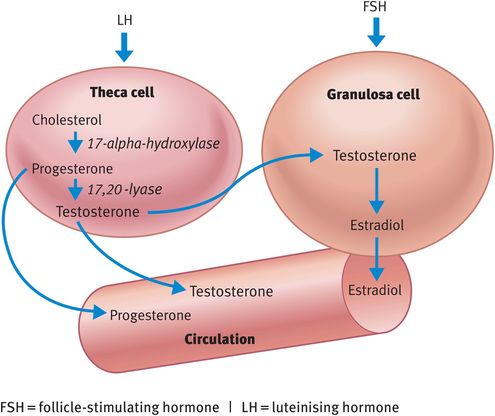
The ovarian steroids are produced in the follicle by sequential loss of carbon atoms from cholesterol (Figure 29.3). LH drives the conversion of cholesterol to androgen by the theca layer. Androgens act as the precursor for estradiol synthesis and FSH primarily drives the conversion of androgen diffusing across the basement membrane to estradiol, a reaction that is catalysed by the enzyme aromatase in the granulosa cells. Both theca and granulosa cells can convert cholesterol into progesterone but only theca cells express CYP 17, which encodes for the enzymes necessary for androgen production, 17-alpha-hydroxylase and 17, 20-lyase, and only granulosa cells contain aromatase. Therefore, both cell types are essential for the production of all steroids. This is known as the two-cell two-gonadotrophin theory. It is the cyclical changes in the peptide hormones and the steroids subsequently produced by the follicles that bring about the menstrual cycle and permit selection of a single ovulatory follicle.
Ovarian steroid production
Assuming that pregnancy did not occur in the previous cycle, the cells in the corpus luteum will die 14 days after ovulation. Death of the cells results in a fall in progesterone and estrogen and subsequent shedding of the endometrium (Figure 29.4). At the beginning of the cycle (day 1) there are from four to six antral follicles of size 2–4 mm in the ovary. As the corpus luteum dies, the fall in steroids results in removal of negative feedback at the hypothalamus and pituitary and this causes a specific rise in FSH (Figure 29.5A). Although both LH and FSH are produced by the pituitary gonadotrophs, it is only FSH that rises at this time. FSH levels remain elevated for approximately 3 days: this is known as the intercycle rise in FSH. It is this rise in FSH that is primarily responsible for selection of the dominant follicle. One follicle has the correct qualities, because of FSH receptor numbers, intracellular cyclic adenosine monophosphate levels or granulosa cell number, and this follicle rapidly increases in size in response. As the number of granulosa cells in the follicle increases exponentially, their products, namely inhibin and estradiol, rise in the serum. This reinstates the negative feedback and FSH levels fall (Figures 29.4 and 29.5B).


The action of FSH on the dominant or selected follicle induces LH receptors on the granulosa cells at this time. No other follicle acquires granulosa cell LH receptors. This means that this follicle can survive the fall in FSH, whereas the other small follicles in the cohort, which are FSH dependent, will die by atresia.
Across the follicular phase, the level of LH gradually increases. The dominant follicle grows from 4 mm to 20 mm with an accompanying considerable increase in granulosa cell number. As each cell is converting androgen from the theca cells into estrogen by the action of aromatase, the serum concentration of estradiol increases rapidly. Once the level has been above 300 nmol/l for 2–3 days, the negative feedback switches to positive.
This causes a rapid and massive release of LH from the pituitary, known as the LH surge (Figures 29.4 and 29.5C).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


