Chapter 440 Development of the Hematopoietic System
Hematopoiesis in the Human Embryo and Fetus
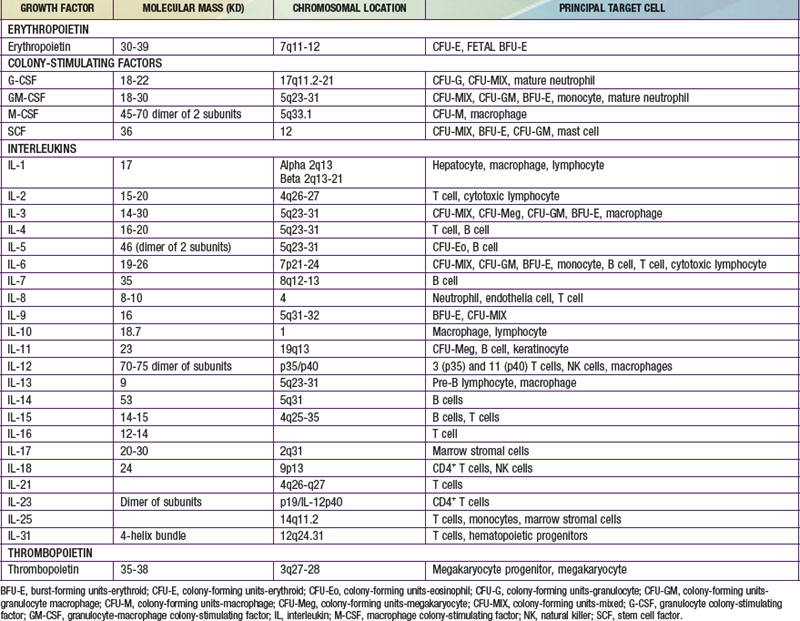
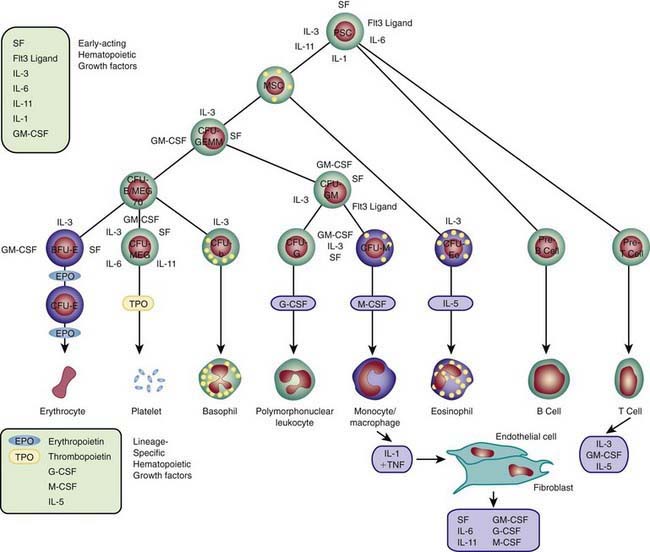
Each hematopoietic organ houses distinct populations of cells. At 18-20 wk, the fetal liver is predominantly an erythropoietic organ, and the marrow produces both erythrocytes and neutrophils. The types of leukocytes present in the fetal liver and marrow differ with gestation. Macrophages precede neutrophils in the marrow, and the ratio of macrophages to neutrophils decreases as gestation progresses. Regardless of gestational age or anatomic location, production of all hematopoietic tissues begins with pluripotent stem cells capable of both self-renewal and clonal maturation into all blood cell lineages. Progenitor cells differentiate under the influence of hematopoietic growth factors (Table 440-1). Fetal hematopoietic growth factor production is independent of maternal growth factor production (Fig. 440-1).
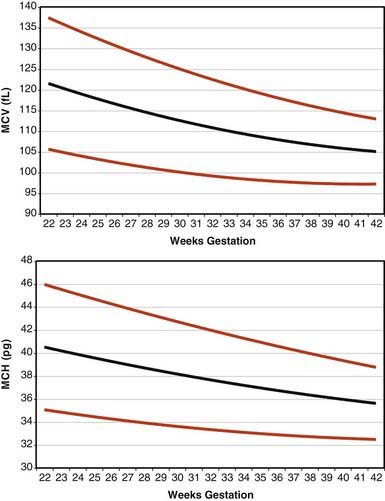
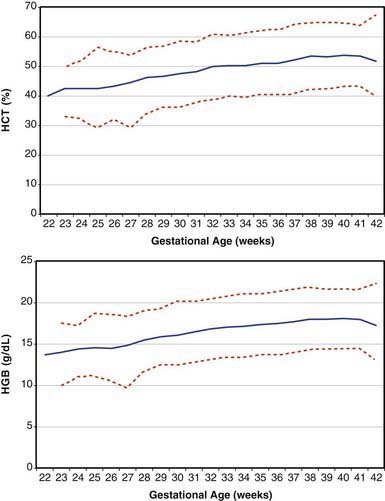
Erythrocytes in the fetus are larger than in adults, and at 22-23 wk gestation the mean corpuscular volume can be as high as 135 fL (Fig. 440-2, upper panel). Similarly the mean corpuscular hemoglobin is very high at 22-23 wk and falls relatively linearly with advancing gestation (see Fig. 440-2, lower panel). In contrast, the mean corpuscular hemoglobin concentration is constant throughout gestation at 34 ± 1 g/fL. Although the size and quantity of hemoglobin in erythrocytes diminish during gestation, the hematocrit and blood hemoglobin concentration gradually increase (Fig. 440-3).
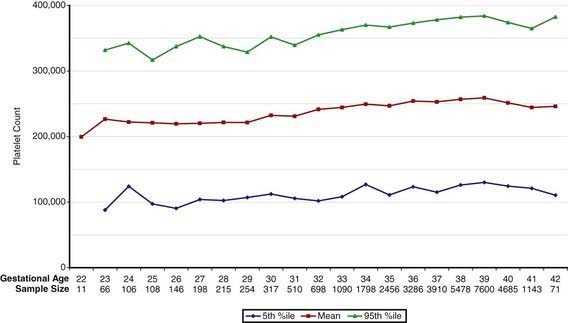
Concentrations of platelets in the blood increase gradually between 22 and 40 wk gestation (Fig. 440-4), but the platelet size, assessed by mean platelet volume, remains constant at 8 ± 1 fL. No differences are observed between males and females in fetal and neonatal reference ranges for erythrocyte indices, hematocrit, hemoglobin, platelet counts, or mean platelet volume measurements.