CHAPTER 10 Hormones
their action and measurement in gynaecological practice
Introduction
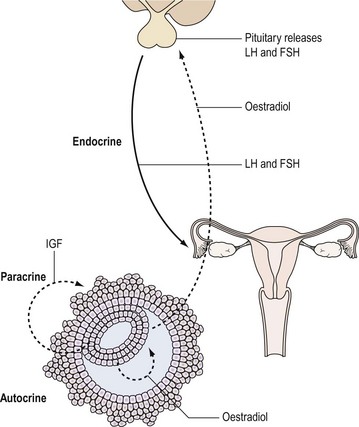
Hormones are one of the important means by which cells communicate with each other. They ensure that the body’s physiological systems are coordinated appropriately, and if this is impaired, as occurs with hormone deficiency or excess, function is compromised. Classically, hormones are synthesized by specialized cells, which may exist as distinct endocrine glands or be located as single cells within other organs, and secreted into the blood stream and transported through the circulation to a distant site of action (endocrine effect). However, autocrine or paracrine actions may also be prevalent and indeed modulate the function of the original endocrine cell type (Figure 10.1).
Hormones: Types and Action
There are three major groups of hormones according to biochemical structure and method of synthesis:
Peptide hormones
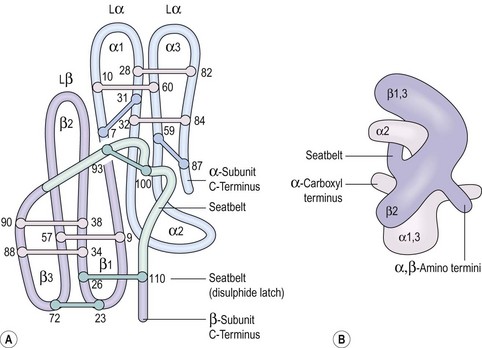
Some peptide hormones have complex tertiary structures; for example, glycoproteins are composed of more than one peptide chain. Oxytocin and arginine vasopressin, two of the posterior pituitary hormones, have ring structures linked by disulphide bridges. Despite being remarkably similar in structure, they have very different physiological roles. The gonadotrophins, follicle-stimulating hormone (FSH) and luteinizing hormone (LH), TSH and human chorionic gonadotrophin (hCG) also have two chains (Figure 10.2). However, these α- and β-subunits are synthesized quite separately from separate genes. The α-subunit of each is shared; it is the distinctive β-subunit of each that confers biological specificity.
Steroids
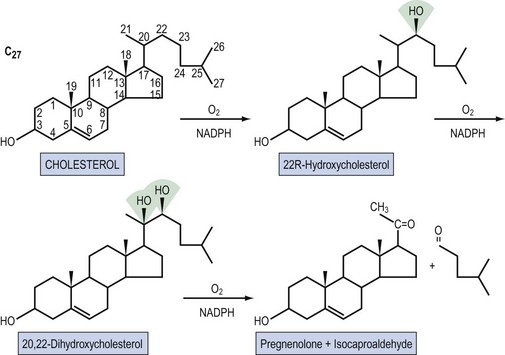
Steroid hormones are lipid-soluble molecules derived from modification of the four-carbon ring structure of cholesterol (Figure 10.3). Steroids can be classified according to the number of carbon atoms they possess: C-21 (progestogens, cortisol, aldosterone), C-19 (androgens) and C-18 (oestrogens). The common names used for steroids also adhere to a loose convention. The suffix -ol indicates an important hydroxyl group, as in cholesterol or cortisol, whereas the suffix -one indicates an important ketone (e.g. testosterone). The extra presence of -di, as in diol (e.g. oestradiol) or dione (e.g. androstenedione), reflects two of these groups, respectively; -ene (androstenedione) within the name indicates a significant double bond in the steroid nucleus.
Regulation of Hormone Secretion and Action
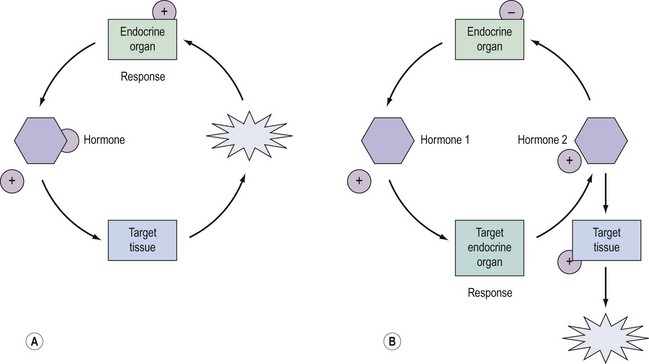
Regulation and control of hormones is essential, and underpins not only the function of many endocrine systems but also their clinical investigation (Figure 10.4). Hormones utilize a variety of differential regulatory mechanisms.
Availability of the hormone to a cell
A hormone can be present in the blood stream but unavailable to the target cell. A hormone circulating in the blood stream, not attached to a binding protein, readily enters cells by simple diffusion. However, most of the steroid hormones circulate bound to protein carriers, such as sex-hormone-binding globulin (SHBG), because their cholesterol origin makes them hydrophobic (King 1988). Consequently, they are generally unavailable, and alterations in the amount of circulating binding globulins can modulate the biological activities of each of their respective hormones. For example, approximately 40% of circulating oestrogen is bound to SHBG, and the remainder is bound to albumin and membrane proteins (Hovarth 1992). As testosterone is largely (80%) bound to SHBG and 19% is loosely bound to albumin, its androgenicity is mainly dependent upon the unbound fraction and partly upon the fraction associated with albumin. SHBG production in the liver is decreased by androgens, hence the binding capacity in men is lower than in normal women. In a hirsute woman, the SHBG level is depressed by the excess androgen, and the percentage of free and therefore active testosterone is elevated (Nestler et al 1991).
Tissue distribution
The tissue distribution of hormone receptors also determines the scope of hormone action, and the presence or absence of a given receptor determines whether a cell will respond to a given class of steroids. For example, the TSH receptor is limited to the thyroid and therefore TSH action is restricted to the thyroid. A hormone can also modify its own and/or another steroid hormone’s activity by regulating the concentration of receptors in a cell. This has the biological effect of increasing tissue response to the hormone if the receptor number is increased, and vice versa if the receptor number is decreased. Oestrogen, for example, increases target tissue responsiveness to itself by increasing the concentration of FSH receptors in granulosa cells (Hillier 1994). This process is important in the selection and maintenance of the dominant ovarian follicle in the menstrual cycle. In order to respond to the ovulatory surge and become a ‘successful’ corpus luteum, the granulosa cells must acquire LH receptors. FSH induces LH receptor development on the granulosa cells of large antral follicles with oestrogen acting as chief coordinator. Progesterone, on the other hand, limits the tissue response to oestrogen by reducing, over time, the concentration of oestrogen receptors, hence its use in the prevention of endometrial hyperplasia and carcinoma (Studd and Wadsworth 2000).
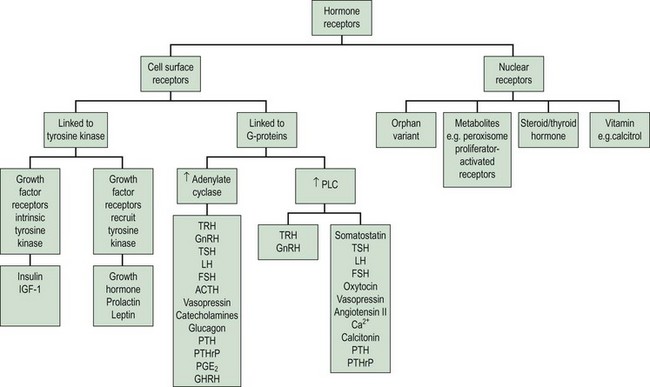
Hormone Receptors
The clinical effects of hormones are mediated through the interaction of a hormone with its receptors. There are two principal superfamilies of hormone receptors. This broad classification is based on their cellular location and their characteristic features (Figure 10.5).
Cell surface receptors
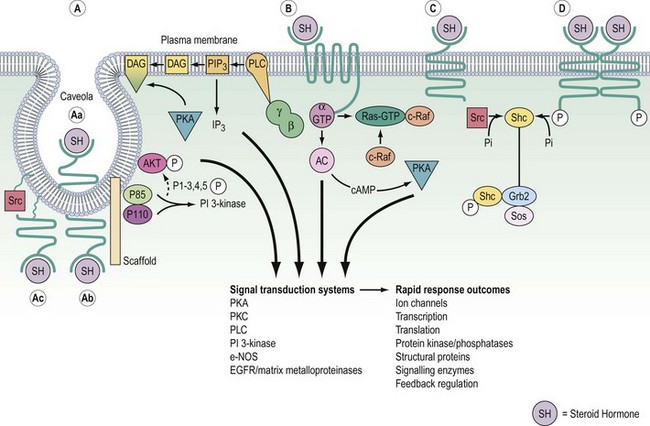
Cell surface receptors are composed of three components, each with structural features, which reflect its location and function (Figure 10.6). The hormone acts as a ligand, with a ligand-binding pocket in the extracellular domain of the receptor being comparatively rich in cysteine residues that form internal disulphide bonds and repeated loops to ensure correct folding. This extracellular domain may also be cleaved for some hormones, and function as a binding protein in the circulation, thus limiting bioavailability (e.g. GH). The α-helical membrane-spanning domain is rich in the hydrophobic and uncharged amino acids, enabling it to be secured with the lipid bilayer. The C-terminal cytoplasmic domain either contains, or links to, separate catalytic systems which initiate the intracellular signals after hormone binding.
Stage 1. Availability of the receptor (up- and downregulation)
The cell’s mechanism for sensing the low concentration of circulating trophic hormone is to have an extremely large number of receptors but to require only a very small percentage (as little as 1%) to be occupied by the hormone for its action to be evident (Sairam and Bhargavi 1985). Positive and negative modulation of receptor numbers by hormones is known as up- and downregulation. The mechanism of upregulation is unclear, but prolactin and GnRH, for example, can increase the concentration of their own receptors in the cell membrane (Katt et al 1985).
In downregulation, an excess concentration of a trophic hormone, such as LH or GnRH, results in a loss of receptors on the cell membranes and therefore a decrease in biological response. This process occurs by internalization of the receptors, and is the main biological mechanism by which the activity of polypeptide hormones is limited (Ron-El et al 2000). Thus the formation of the hormone–receptor complex on the cell surface initiates the cellular response, and the internalization of the complex (with eventual degradation of the hormone) terminates the response. It therefore appears that the principal reason for the pulsatile secretion of trophic hormones is to avoid downregulation and to maintain adequate receptor numbers. The pulse frequency, therefore, is a key factor in regulating receptor number.
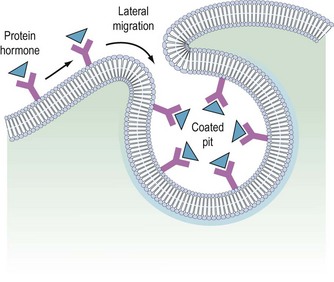
It is believed that receptors are randomly inserted into the cell membrane after intracellular synthesis. They have two important sites — an external binding site, which is specific for a polypeptide hormone, and an internal site, which plays a role in the process of internalization (Kaplan 1981). When a hormone binds to the receptor and a high concentration of the hormone is present in the circulation, the hormone–receptor complex moves laterally in the cell membrane by a process called ‘lateral migration’ to a specialized area, the coated pit, the internal margin of which has a brush border (Goldstein et al 1979). Lateral migration, which takes minutes rather than seconds, thus concentrates hormone–receptor complexes in the coated pit, a process referred to as ‘clustering’ (Figure 10.7). When fully occupied, the coated pit invaginates, ‘pinches off’ and enters the cell as a vesicle. The coated vesicle is delivered to the lysosomes where it undergoes degradation, releasing the hormone receptors. The receptors may be recycled to the cell membranes and used again, or the receptor and hormone may be metabolized, thus decreasing the hormone’s biological activity. This process is called ‘receptor-mediated endocytosis’ (Goldstein et al 1979; Figure 10.8).
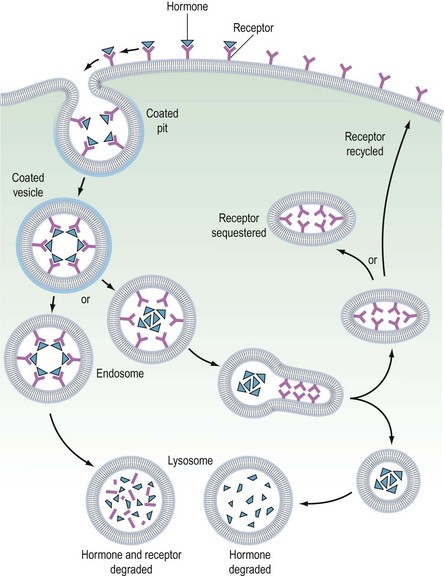
As well as downregulation of polypeptide hormone receptors, the process of internalization can be utilized for other cellular metabolic events, including the transfer of vital substances into the cell, such as iron and vitamins. Hence, cell membrane receptors can be separated into two classes. Class I receptors are distributed in the cell membrane and transmit information to modify cell behaviour for these receptors. Internalization is a method for downregulation and recycling is not usually a feature. Hormones which utilize this category of receptor include FSH, LH, hCG, GnRH, TSH and insulin (Kaplan 1981). Class II receptors are located in the coated pits. Binding leads to internalization, which provides the cell with the required ligands or removes noxious agents from the biological fluid bathing the cell. These receptors are spared from degradation and can be recycled. Examples of this category include low-density lipoproteins, which supply cholesterol to steroid-producing cells (Parinaud et al 1987), and transfer of immunoglobulins across the placenta to provide fetal immunity.
Stage 2. Binding of the hormone to the receptor
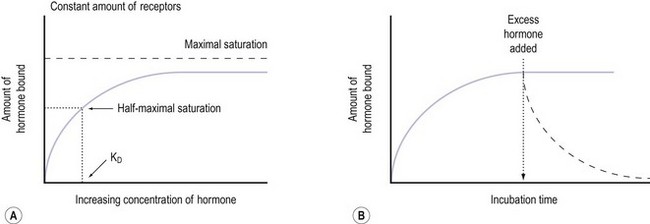
Notably hormone–receptor systems demonstrate standard kinetics and are saturable and reversible (Figure 10.9). Consequently, the dissociation rate of the hormone and its receptor is an important component of the biological response, as the biological activity of the hormone is only maintained while the hormone–receptor complex is maintained. Hormones that are structurally similar may have some overlap in biological activity. For example, the similarity in structure between GH and prolactin means that GH has a lactogenic action whilst prolactin has some growth-promoting activity and stimulates somatomedin production.
The glycoprotein hormones (LH, FSH, TSH and hCG) share an identical α-chain and require another portion, the β-chain, to confer the specificity inherent in the relationship between hormones and their receptors. The β-subunits differ in both amino acid and carbohydrate content, and the chemical composition may be altered under certain conditions, thereby affecting the affinity of the hormone and its receptor (Willey 1999).
Stage 3. Signal transduction
On binding of a hormone to its cell surface receptor, protein phosphorylation and the generation of ‘second messengers’ mediates a cascade of cytoplasmic and cellular responses. This multistep cascade facilitates amplification of the cellular response to the hormone binding to a receptor, as many second messenger molecules are produced for each hormone–receptor complex. The cell surface receptors may be classified into two groups according to the second messenger and signalling cascades that are stimulated: (1) tyrosine kinase receptors and (2) G-protein coupled receptors (Figure 10.5).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


