CHAPTER 11 Biosynthesis of steroid hormones
Physiology
The classification of steroid hormones
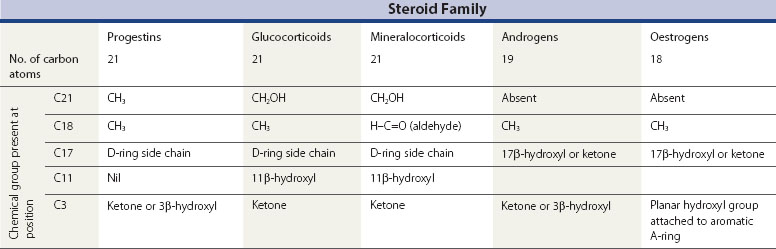
All steroid hormones are ultimately derived from the 27 carbon (C) substrate cholesterol and so share the same cyclohexaphenanthrene ring structure (Figures 11.1 and 11.2). Steroids are classified into five families dependent upon the number of carbon atoms and the chemical groups present at key carbon residues (Table 11.1). Those steroid hormones with 21 carbon atoms are collectively termed ‘pregnenes’, and this category of steroid hormones can be subdivided into three steroid families: progestins (e.g. progesterone), glucocorticoids (e.g. cortisol) and mineralocorticoids (e.g. aldosterone). While the progestins are secreted predominantly from the ovary, the glucocorticoids and mineralocorticoids are collectively termed ‘corticosteroids’, reflecting their origin in the cortex of the adrenal gland. The major structural difference between the three pregnene families is that while progestins possess a methyl group (CH3) at position C21, both glucocorticoids and mineralocorticoids possess a C21 hydroxyl group (CH2OH) (see Table 11.1 and Figure 11.2). Progestins can be metabolized to generate 19 carbon steroids, termed ‘androgens’ [dehydroepiandrosterone (DHEA), androstenedione and testosterone], which are secreted from both the testis and the adrenal cortex. Within the ovary, androgens are usually metabolized to generate the oestrogens (e.g. oestradiol-17β) with their characteristic 18 carbon structure (see Table 11.1 and Figure 11.2).
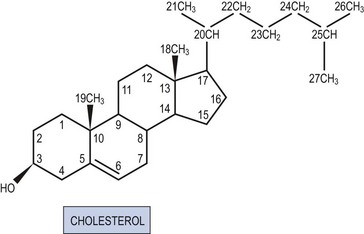
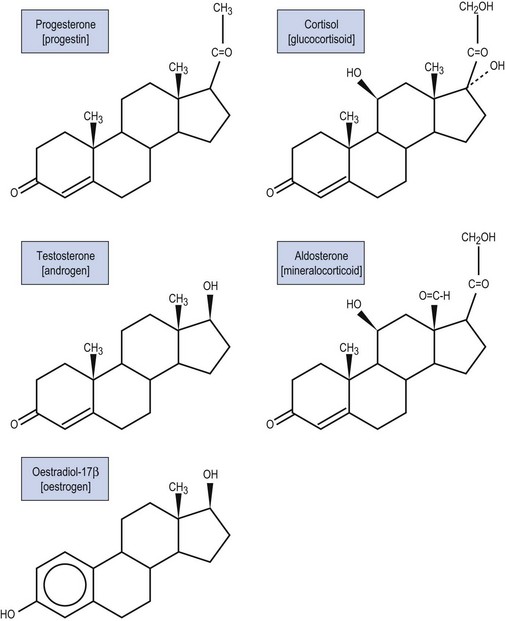
Within this classification scheme, there is a distinction to be made between Δ5 and Δ4 steroid hormones. Progestins and androgens of the Δ5 series are characterized by possessing a hydroxyl group at position C3 and a C C double bond between positions C5 and C6 in the steroid B-ring, as in cholesterol (Figure 11.1). In contrast, progestins and androgens of the Δ4 series possess a ketone (C
C double bond between positions C5 and C6 in the steroid B-ring, as in cholesterol (Figure 11.1). In contrast, progestins and androgens of the Δ4 series possess a ketone (C O) at position C3 and have their C
O) at position C3 and have their C C double bond between positions C4 and C5 in the A-ring of the steroid molecule (Figure 11.2). While this difference may seem a trivial biochemical detail, nothing could be further from the truth. The nature of the chemical group at position C3, together with the position of the C
C double bond between positions C4 and C5 in the A-ring of the steroid molecule (Figure 11.2). While this difference may seem a trivial biochemical detail, nothing could be further from the truth. The nature of the chemical group at position C3, together with the position of the C C double bond, profoundly alters the conformation of the steroid molecule and, in so doing, influences the ability of the hormone to activate intracellular receptors. Hence, Δ5 steroids, such as pregnenolone and DHEA, have low affinities for steroid receptors such that they can only exert limited biological actions. In contrast, the Δ4 steroids, such as progesterone and testosterone, are potent activators of steroid receptors and so act as the dominant physiological hormones.
C double bond, profoundly alters the conformation of the steroid molecule and, in so doing, influences the ability of the hormone to activate intracellular receptors. Hence, Δ5 steroids, such as pregnenolone and DHEA, have low affinities for steroid receptors such that they can only exert limited biological actions. In contrast, the Δ4 steroids, such as progesterone and testosterone, are potent activators of steroid receptors and so act as the dominant physiological hormones.
Origins and intracellular transport of cholesterol
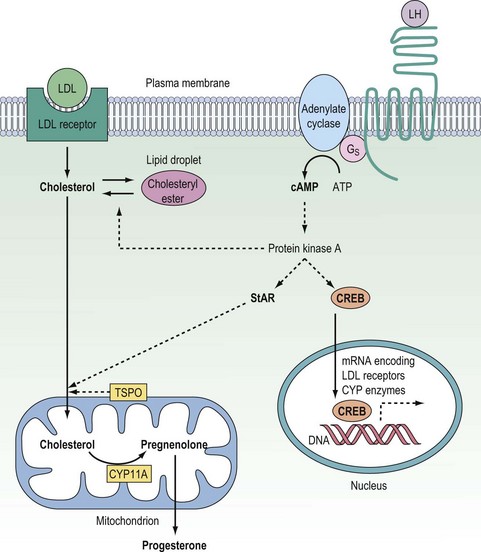
Once inside the cell, cholesterol must be transported across the cytosol and into the mitochondria where the first and rate-determining reaction of steroid synthesis occurs: the catabolism of cholesterol to form pregnenolone (see below). The transport of cholesterol across the two mitochondrial membranes and interceding aqueous intermembrane space is now recognized as the true rate-limiting step for steroidogenesis, and so with this realization, it is this process which has been the focus of research into steroid biosynthesis over the past two decades. Although the understanding of the molecular mechanisms by which cholesterol passes from the outer leaflet of the outer mitochondrial membrane (OMM) to the inner leaflet of the inner mitochondrial membrane (IMM) is still far from complete, significant advances have been made over the past decade. In the mid 1990s, the importance of the steroidogenic acute regulatory (StAR) protein was first indicated when Clark and Stocco showed StAR to be a short half-life protein which is rapidly upregulated in steroidogenic cells following stimulation either by trophic hormones (e.g. gonadotrophins) or by the common second messenger, cyclic adenosine 3′,5′-monophosphate (cAMP) (Stocco and Clark 1996). Analysis of the StAR peptide sequence suggested that this protein gets imported from the cytosol into the IMM, and it was postulated that the StAR protein might simply bind and transport cholesterol across the aqueous intermembrane space during the course of that import. However, subsequent site-directed mutagenesis studies revealed that the StAR protein could still drive cholesterol flux for steroidogenesis even when it was modified to prevent mitochondrial import, and it is now accepted that the StAR protein can act at the OMM to drive cholesterol passage from the outer leaflet of the OMM to the inner leaflet of the IMM without itself trafficking into the mitochondria (Miller 2007). This steroidogenic action of the StAR protein does, however, require: (i) that the StAR protein be phosphorylated on key serine residues by the cAMP-dependent protein kinase A (PKA); and (ii) that the phosphorylated form of the StAR protein interacts with the 18 kDa translocator protein (TSPO) expressed in the OMM (Figure 11.3) (Hauet et al 2005, Miller 2007). The TSPO protein, which was originally described as a peripheral benzodiazepine receptor (PBR), is very highly expressed in the OMM of steroidogenic cells. Recent studies have revealed that rather than reliance on a single protein to drive mitochondrial cholesterol uptake for steroidogenesis, this pivotal rate-limiting step involves a complex of proteins which includes the StAR and TSPO proteins in association with PBR-associated protein-7 (PAP7). By virtue of its structural interactions with both the TSPO protein and the PKAR1α regulatory subunit of PKA, PAP7 acts as a PKA-anchoring protein, localizing PKA to the OMM where it can phosphorylate the serine residues in the StAR protein to drive the StAR–TSPO interaction required to deliver cholesterol to the IMM for the first reaction in the steroidogenic pathway (Liu et al 2006, Miller 2007). While the StAR protein is not expressed in the placenta, this tissue does express a homologous protein, metastatic lymph node 64, which shares several functional and structural properties with the StAR protein, and is assumed to drive mitochondrial cholesterol uptake for placental steroidogenesis (Petrescu et al 2001).
Steroidogenic enzymes
Although the biochemical pathway of steroid hormone synthesis, summarized in Figure 11.4, can look like a bewildering railway map, the whole pathway becomes more straightforward on recognizing that the eight major steroidogenic enzymes can all be classified into just two enzyme families (the cytochrome P450 enzymes and the hydroxysteroid dehydrogenases), each with definitive functions. The steroidogenic cytochrome P450 (CYP) enzymes are members of a larger superfamily of CYP oxidase enzymes. Each CYP enzyme catalyses a hydroxylation reaction by serving as the terminal electron acceptor in an electron transport pathway which transfers electrons from a pyridine nucleotide cofactor, nicotinamide adenine dinucleotide phosphate, via two sequential flavoproteins (ferrodoxin reductase and ferrodoxin) on to atomic oxygen (Miller 2008). This allows the atomic oxygen to be inserted, by the CYP enzyme, into a hydrocarbon (C․H) bond thus generating a polar hydroxyl group (C․O․H) which renders the substrate molecule more water soluble/hydrophilic. The non-steroidogenic CYP enzymes include those hepatic oxidase enzymes responsible for the inactivation and clearance of a wide range of drugs. Each of the CYP enzymes within the steroidogenic pathway has traditionally been referred to by a variety of names that reflect their ability to catalyse hydroxylation of a steroid substrate at a specific carbon position. However, following the completion of the human genome (HUGO) project, all steroidogenic CYP enzymes have been assigned new systematic names which relate to the CYP gene that encodes the enzyme protein (Table 11.2).
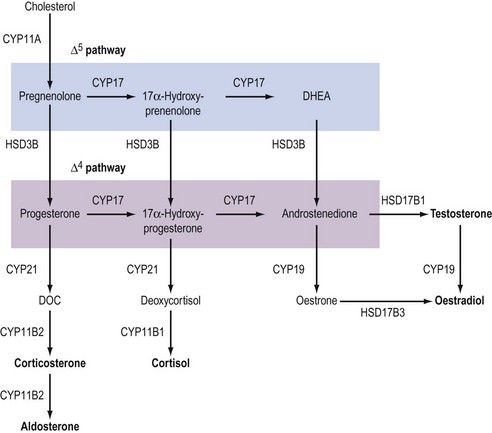
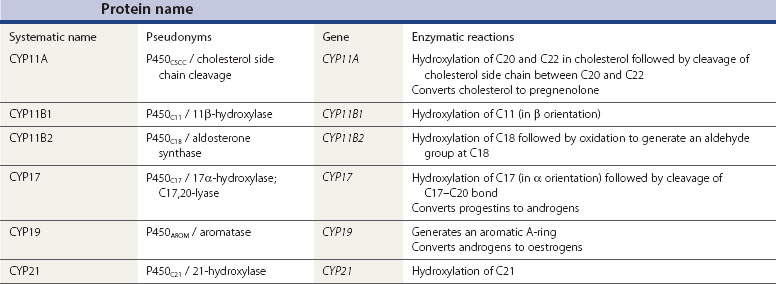
In addition to their characteristic hydroxylase activities, some steroidogenic CYP enzymes also catalyse lyase reactions. Specifically, the CYP11A (cytochrome P450 cholesterol side chain cleavage) and CYP17 (cytochrome P450 17α-hydroxylase/C17,20-lyase) enzymes can each catalyse cleavage of the C․C bond weakened by the hydroxylation reactions. CYP11A acts in the IMM to catalyse the rate-limiting reaction in steroid synthesis: the conversion of cholesterol to pregnenolone. This crucial CYP enzyme hydroxylates two adjacent carbons (C20 and C22) in the D-ring side chain of cholesterol, facilitating cleavage between C20 and C22 to leave the Δ5 21 carbon steroid, pregnenolone (Miller 2008). Similarly, CYP17 introduces a hydroxyl group at position C17 of either pregnenolone or progesterone, as a result of which the weakened C17–C20 bond breaks to generate either DHEA or androstenedione, respectively (Miller 2008) (see Figure 11.4). Other members of the CYP enzyme family simply catalyse introduction of oxygen to generate hydroxyl groups at specific carbon positions (see Table 11.2). The hydroxylations catalysed by CYP21 (21-hydroxylase) and CYP11B1 (11β-hydroxylase) are pivotal in the formation of corticosteroids by the adrenal cortex (Miller 2008). The hydroxyl group generated at position C18 by CYP11B2 (aldosterone synthase) undergoes rapid oxidation to form an aldehyde group (Curnow et al 1991), hence giving rise to the name ‘aldosterone’ (see Figure 11.2). Finally, CYP19 (aromatase) is the most complex member of the steroidogenic CYP enzyme family, catalysing a series of reactions that convert C19 androgens (androstenedione and testosterone) to their C18 oestrogen metabolites (oestrone and oestradiol, respectively). In this reaction sequence, the C19 methyl group is lost and the ketone at position C3 is reduced to a hydroxyl group (Miller 2008). This liberates electrons which are invested in the A-ring of the steroid to generate the aromatic phenol ring, the hallmark of oestrogens (see Figure 11.2) and a prerequisite for activation of the oestrogen receptor.
The second family of steroidogenic enzymes comprises the hydroxysteroid dehydrogenase (HSD) enzymes encoded by HSD genes (Penning 1997, Miller 2008). (As with the CYP enzymes, the HSD enzymes have all been renamed following the completion of the HUGO project, such that the systematic names for the 3βHSD and 17βHSD isoenzymes have changed to HSD3B and HSD17B, respectively.) All of the HSD enzymes catalyse the interconversion of a hydroxyl group with a ketone to influence the affinity of the steroid hormone for its intracellular receptors (Penning 1997). Each HSD enzyme is specified by a number and a Greek letter; the number denotes the position of the carbon atom at which the enzyme acts, and the Greek letter indicates the orientation of the OH group relative to the steroid molecule (where α and β indicate bonds below and above the plane of the molecule, respectively).
There are two HSD enzymes of significance in steroid hormone production. The two cloned HSD3B isoenzymes convert the 3β-hydroxyl group of weak Δ5 steroids to a ketone, and also catalyse movement of the C C double bond to generate the corresponding Δ4 steroid metabolite, thus increasing steroid potency (Penning 1997, Miller 2008). Hence, both the placental HSD3B1 enzyme and HSD3B2, expressed in the ovary, testis and adrenal gland, can convert weak Δ5 steroids, such as pregnenolone and DHEA, to the active Δ4 progestins and androgens, progesterone and androstenedione, respectively (see Figure 11.4). In contrast, the major HSD17B isoenzymes act at position C17 where they catalyse not oxidation but reduction, converting a ketone at C17 to a β-hydroxyl group. The significance of this reaction is attested to by the fact that androstenedione and oestrone (which have ketone groups at position C17) are relatively weak steroids, whereas their 17β-hydroxy-metabolites, testosterone and oestradiol-17β (see Figure 11.2), are the more potent androgen and oestrogen, respectively.
C double bond to generate the corresponding Δ4 steroid metabolite, thus increasing steroid potency (Penning 1997, Miller 2008). Hence, both the placental HSD3B1 enzyme and HSD3B2, expressed in the ovary, testis and adrenal gland, can convert weak Δ5 steroids, such as pregnenolone and DHEA, to the active Δ4 progestins and androgens, progesterone and androstenedione, respectively (see Figure 11.4). In contrast, the major HSD17B isoenzymes act at position C17 where they catalyse not oxidation but reduction, converting a ketone at C17 to a β-hydroxyl group. The significance of this reaction is attested to by the fact that androstenedione and oestrone (which have ketone groups at position C17) are relatively weak steroids, whereas their 17β-hydroxy-metabolites, testosterone and oestradiol-17β (see Figure 11.2), are the more potent androgen and oestrogen, respectively.
Regulation of Steroid Synthesis
In the ovary, testis and adrenal cortex, the synthesis of steroid hormones is regulated primarily by trophic hormones secreted from the anterior lobe of the pituitary gland, acting in conjunction with other endocrine, paracrine and autocrine modulators of steroidogenesis. The anterior pituitary hormones, being hydrophilic, have to act via cell surface receptors coupled to signal transduction pathways that increase the expression and activities of steroidogenic enzymes. While several second messengers are generated in response to trophic hormones, the steroidogenic responses to these proteins are mediated primarily through the generation of cAMP with activation of PKA. In the acute response to endocrine stimulation, activation of PKA phosphorylates: (i) cholesteryl ester hydrolase (increasing mobilization of cholesterol from intracellular lipid droplets); and (ii) the StAR protein (thus promoting mitochondrial uptake of cholesterol for steroid biosynthesis) (Niswender 2002, Miller 2008). In addition, activation of the cAMP–PKA system rapidly upregulates expression of the StAR protein, apparently via the cAMP-response element modulator (CREM) protein interacting with a non-classical cAMP response element half-site in the promoter region of the StAR gene (Sugawara et al 2006
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


